Difference between revisions of "Amino acid" - New World Encyclopedia
| Line 41: | Line 41: | ||
=== Nonstandard amino acids === | === Nonstandard amino acids === | ||
| − | Aside from the twenty standard amino acids and | + | Aside from the twenty standard amino acids and two special amino acids, selenocysteine and pyrrolysine, which are coded by DNA in an atypical manner, there are a vast number of [[nonstandard amino acids]]. These nonstandard or ''non-proteinogenic amino acids'' are either not found in proteins (like [[carnitine]], [[GABA]], or [[L-DOPA]]), or not coded for in the standard genetic code (like hydroxyproline and selenomethionine). The latter often result from [[posttranslational modification]] of proteins: the modification of standard amino acids after the protein has been formed in the translation stage of protein synthesis. |
| − | + | Some of these non-standard amino acids have been detected in [[meteorite]]s, especially in a type known as [[carbonaceous chondrite]]s, whose composition is considered to be representative of the [[solar nebula]], or gaseous cloud, from which the solar system condensed. For example, over 79 amino acids were found in the primitive [[Murchison meteorite]], a type of carbonaceous chondrite. [[Microorganism]]s and [[plant]]s can also produce uncommon amino acids that can be found in peptidic [[antibiotics]] such as [[nisin]], which is used as a food preservative. | |
==The structure of amino acids== | ==The structure of amino acids== | ||
| − | In [[biochemistry]], the term "amino acid" is frequently used to refer specifically to ''alpha amino acids'': those amino acids in which the amino and carboxylate groups are attached to the same [[carbon]], the so-called [[alpha carbon|α–carbon]]. The general structure of proteinogenic alpha amino acids is: | + | In [[biochemistry]], the term "amino acid" is frequently used to refer specifically to ''alpha amino acids'': those amino acids in which the amino and carboxylate groups are attached to the same [[carbon]], the so-called [[alpha carbon|α–carbon]]. The general structure of these proteinogenic alpha amino acids is: |
''R'' | ''R'' | ||
| Line 53: | Line 53: | ||
| | | | ||
H | H | ||
| − | where ''R'' represents a ''side chain'' specific to each amino acid. The exception is [[proline]], whose side chain cyclizes onto the backbone | + | where ''R'' represents a ''side chain'' specific to each amino acid. The exception to this basic structure is [[proline]], whose side chain cyclizes onto the backbone, forming a ring structure in which a secondary amino group replaces the primary amino group. |
| − | When amino acids join to form proteins, the basic amino group and acidic carboxyl group are neutralized | + | When amino acids join together to form proteins, the basic amino group and acidic carboxyl group are neutralized. Instead, the side chains of the amino acids determine the acid-base properties of proteins. Thus, amino acids are usually classified by two main [[chemical properties]] of the side chain: the charge of the R group (which determines whether the protein acts like a weak acid or a weak base), and its polarity (or tendency to interact with water at a neutral pH). |
| − | These properties influence the amino acids' interaction with other structures | + | These properties influence the amino acids' interaction with other structures within the protein as well as their interaction with other proteins. For example, soluble proteins have surfaces rich with polar amino acids like serine and threonine. In contrast, [[integral membrane protein]]s tend to have outer ring of [[hydrophobic]] amino acids (which do not tend to interact with water) that anchors them to the [[lipid bilayer]]. Similarly, proteins that have to bind to positively charged molecules have surfaces rich with negatively charged amino acids like glutamate and aspartate, while proteins binding to negatively charged molecules have surfaces rich with positively charged chains like lysine and arginine. |
===Isomers=== | ===Isomers=== | ||
| Line 64: | Line 64: | ||
==Amino acids function in protein synthesis== | ==Amino acids function in protein synthesis== | ||
[[Image:Peptide_bonding.gif|thumb|Summary of the formation of a peptide bond.]] | [[Image:Peptide_bonding.gif|thumb|Summary of the formation of a peptide bond.]] | ||
| − | Two amino acids are linked together by a peptide bond, which | + | Two amino acids are linked together by a [[peptide bond]], which forms when the basic amino group of one amino acid reacts with the acidic carboxyl group of a second amino acid. This [[condensation reaction]] (water loss) yields a peptide bond and a molecule of water. An '''amino acid residue''' is what is left of an amino acid once a molecule of [[water]] has been lost (an [[hydrogen ion|H<sup>+</sup>]] from the amino group side and an [[hydroxyl ion|OH<sup>-</sup>]] from the carboxylic side) in the formation of a peptide bond. |
Proteins are then created by the [[polymerization]] of amino acids, a process in which amino acids are joined together in chains called, depending on their length, [[peptides]] or [[polypeptides]]. | Proteins are then created by the [[polymerization]] of amino acids, a process in which amino acids are joined together in chains called, depending on their length, [[peptides]] or [[polypeptides]]. | ||
==Other biological roles of amino acids== | ==Other biological roles of amino acids== | ||
| − | In addition to | + | In addition to their function in the formation of proteins, amino acids have other biologically-important roles. The standard amino acids [[glycine]] and [[glutamate]], which are found in proteins, also function as [[neurotransmitter]]s that respectively inhibit and amplify signals in the nervous system. The non-standard amino acid [[carnitine]] is used in [[lipid]] transport within the [[cell (biology)|cell]]. |
Many amino acids are used to synthesize molecules other than proteins: | Many amino acids are used to synthesize molecules other than proteins: | ||
| Line 79: | Line 79: | ||
*[[Aspartame]] (aspartyl-phenylalanine-1-methyl ester) is an artificial sweetener, and [[monosodium glutamate]] is a [[food additive]] that enhances flavor. | *[[Aspartame]] (aspartyl-phenylalanine-1-methyl ester) is an artificial sweetener, and [[monosodium glutamate]] is a [[food additive]] that enhances flavor. | ||
*[[5-HTP]] (5-hydroxytryptophan) has been used to treat neurological problems associated with [[PKU]] (phenylketonuria), as well as depression (as an alternative to L-Tryptophan). | *[[5-HTP]] (5-hydroxytryptophan) has been used to treat neurological problems associated with [[PKU]] (phenylketonuria), as well as depression (as an alternative to L-Tryptophan). | ||
| − | *[[L-DOPA]] (L-dihydroxyphenylalanine) is a drug used to treat the symptoms of [[Parkinson's disease]] | + | *[[L-DOPA]] (L-dihydroxyphenylalanine) is a drug used to treat the symptoms of [[Parkinson's disease]]. |
==Table of chemical properties== | ==Table of chemical properties== | ||
| − | Following is a table listing the | + | Following is a table listing the symbols and chemical properties of the standard amino acids. The mass listed is the weighted average of all common isotopes, and includes the mass of H<sub>2</sub>O. The one-letter symbol for an undetermined amino acid is ''X''. The three-letter symbol ''Asx'' or one-letter symbol ''B'' means the amino acid is either [[asparagine]] or [[aspartic acid]], whereas ''Glx'' or ''Z'' means either [[glutamic acid]] or [[glutamine]]. The three-letter symbol ''Sec'' or one-letter symbol ''U'' refers to [[selenocysteine]]. The letters ''J'' and ''O'' are not used. |
Essential amino acids are marked with an asterisk. | Essential amino acids are marked with an asterisk. | ||
| Line 108: | Line 108: | ||
| 9.87 | | 9.87 | ||
| | | | ||
| − | | Very abundant | + | | Very abundant and versatile. It behaves fairly neutrally and can be located in both hydrophilic regions on the outside of the protein and hydrophobic interior areas. |
|- | |- | ||
| C | | C | ||
| Line 119: | Line 119: | ||
| 10.70 | | 10.70 | ||
| 8.18 | | 8.18 | ||
| − | | The sulfur atom binds readily to [[heavy metals|heavy metal]] ions. Under oxidizing conditions, two cysteines can | + | | The sulfur atom binds readily to [[heavy metals|heavy metal]] ions. Under oxidizing conditions, two cysteines can be joined together by a [[disulfide bond]] to form the amino acid [[cystine]]. When cystines are components of a protein, they enforce [[tertiary structure]] and makes the protein more resistant to unfolding and [[denaturation (biochemistry)|denaturation]]; disulphide bridges are therefore common in proteins that have to function in harsh environments, digestive enzymes (e.g., [[pepsin]] and [[chymotrypsin]]), structural proteins (e.g., [[keratin]]), and proteins too small to hold their shape on their own (eg. [[insulin]]). |
|- | |- | ||
| D | | D | ||
| Line 130: | Line 130: | ||
| 9.90 | | 9.90 | ||
| 3.90 | | 3.90 | ||
| − | | Behaves similarly to glutamic acid. Carries a hydrophilic acidic group with strong negative charge. Usually is located on the outer surface of the protein, making it water-soluble. Binds to positively-charged molecules and ions, often used in enzymes to fix the metal ion | + | | Behaves similarly to glutamic acid. Carries a hydrophilic acidic group with strong negative charge. Usually is located on the outer surface of the protein, making it water-soluble. Binds to positively-charged molecules and ions, often used in enzymes to fix the metal ion. |
|- | |- | ||
| E | | E | ||
| Line 143: | Line 143: | ||
| Behaves similar to aspartic acid. Has longer, slightly more flexible side chain. | | Behaves similar to aspartic acid. Has longer, slightly more flexible side chain. | ||
|- | |- | ||
| − | | F | + | | *F |
| Phe | | Phe | ||
| − | | [[Phenylalanine]] | + | | [[Phenylalanine]] |
| hydrophobic | | hydrophobic | ||
| 165.19 | | 165.19 | ||
| Line 152: | Line 152: | ||
| 9.31 | | 9.31 | ||
| | | | ||
| − | | Phenylalanine, tyrosine, and tryptophan contain large rigid [[aromaticity|aromatic]] group on the side chain. These are the biggest amino acids. Like isoleucine, leucine and valine, | + | | Phenylalanine, tyrosine, and tryptophan contain large rigid [[aromaticity|aromatic]] group on the side chain. These are the biggest amino acids. Like isoleucine, leucine and valine, they are hydrophobic and tend to orient towards the interior of the folded protein molecule. |
|- | |- | ||
| G | | G | ||
| Line 163: | Line 163: | ||
| 9.78 | | 9.78 | ||
| | | | ||
| − | | Because of the two hydrogen atoms at the α carbon, glycine is not [[optical isomerism|optically active]]. It is the smallest amino acid, rotates easily, adds flexibility to the protein chain. It is able to fit into the tightest spaces, e.g., the triple helix of [[collagen]] | + | | Because of the two hydrogen atoms at the α carbon, glycine is not [[optical isomerism|optically active]]. It is the smallest amino acid, rotates easily, and adds flexibility to the protein chain. It is able to fit into the tightest spaces, e.g., the triple helix of [[collagen]]. |
|- | |- | ||
| H | | H | ||
| His | | His | ||
| − | | [[Histidine]] | + | | [[Histidine]] |
| [[basic (chemistry)|basic]] | | [[basic (chemistry)|basic]] | ||
| 155.16 | | 155.16 | ||
| Line 174: | Line 174: | ||
| 9.33 | | 9.33 | ||
| 6.04 | | 6.04 | ||
| − | |In even slightly acidic conditions, [[protonation]] of the nitrogen occurs, changing the properties of histidine and the polypeptide as a whole. It is used by many proteins as a regulatory mechanism, changing the conformation and behavior of the polypeptide in acidic regions such as the late [[endosome]] or [[lysosome]], enforcing conformation change in enzymes | + | |In even slightly acidic conditions, [[protonation]] of the nitrogen occurs, changing the properties of histidine and the polypeptide as a whole. It is used by many proteins as a regulatory mechanism, changing the conformation and behavior of the polypeptide in acidic regions such as the late [[endosome]] or [[lysosome]], enforcing conformation change in enzymes. |
|- | |- | ||
| − | | I | + | | *I |
| Ile | | Ile | ||
| [[Isoleucine]] | | [[Isoleucine]] | ||
| Line 187: | Line 187: | ||
| Isoleucine, leucine and valine have large aliphatic hydrophobic side chains. Their molecules are rigid, and their mutual hydrophobic interactions are important for the correct folding of proteins, as these chains tend to be located inside of the protein molecule. | | Isoleucine, leucine and valine have large aliphatic hydrophobic side chains. Their molecules are rigid, and their mutual hydrophobic interactions are important for the correct folding of proteins, as these chains tend to be located inside of the protein molecule. | ||
|- | |- | ||
| − | | K | + | | *K |
| Lys | | Lys | ||
| − | | [[Lysine]] | + | | [[Lysine]] |
| basic | | basic | ||
| 146.19 | | 146.19 | ||
| Line 196: | Line 196: | ||
| 9.06 | | 9.06 | ||
| 10.54 | | 10.54 | ||
| − | | Behaves similarly to arginine. Contains a long flexible side-chain with a positively-charged end. The flexibility of the chain makes lysine and arginine suitable for binding to molecules with many negative charges on their surfaces. E.g., [[deoxyribonucleic acid|DNA]]-binding proteins have their active regions rich with arginine and lysine. The strong charge makes these two amino acids prone to be located on the outer hydrophilic surfaces of the proteins | + | | Behaves similarly to arginine. Contains a long flexible side-chain with a positively-charged end. The flexibility of the chain makes lysine and arginine suitable for binding to molecules with many negative charges on their surfaces. E.g., [[deoxyribonucleic acid|DNA]]-binding proteins have their active regions rich with arginine and lysine. The strong charge makes these two amino acids prone to be located on the outer hydrophilic surfaces of the proteins. |
|- | |- | ||
| − | | L | + | | *L |
| Leu | | Leu | ||
| − | | [[Leucine]] | + | | [[Leucine]] |
| hydrophobic | | hydrophobic | ||
| 131.17 | | 131.17 | ||
| Line 209: | Line 209: | ||
| Behaves similar to isoleucine and valine. See isoleucine. | | Behaves similar to isoleucine and valine. See isoleucine. | ||
|- | |- | ||
| − | | M | + | | *M |
| Met | | Met | ||
| − | | [[Methionine]] | + | | [[Methionine]] |
| hydrophobic | | hydrophobic | ||
| 149.21 | | 149.21 | ||
| Line 218: | Line 218: | ||
| 9.28 | | 9.28 | ||
| | | | ||
| − | | Always the first amino acid to be incorporated into a protein; sometimes removed after translation. Like cysteine, contains sulfur, but with a [[methyl]] group instead of hydrogen. This methyl group can be activated, and is used in many reactions where a new carbon atom is being added to another molecule. | + | | Always the first amino acid to be incorporated into a protein; sometimes removed after translation. Like cysteine, it contains sulfur, but with a [[methyl]] group instead of hydrogen. This methyl group can be activated, and is used in many reactions where a new carbon atom is being added to another molecule. |
|- | |- | ||
| N | | N | ||
| Line 275: | Line 275: | ||
| Serine and threonine have a short group ended with a [[hydroxyl]] group. Its hydrogen is easy to remove, so serine and threonine often act as hydrogen donors in enzymes. Both are very hydrophilic, therefore the outer regions of soluble proteins tend to be rich with them. | | Serine and threonine have a short group ended with a [[hydroxyl]] group. Its hydrogen is easy to remove, so serine and threonine often act as hydrogen donors in enzymes. Both are very hydrophilic, therefore the outer regions of soluble proteins tend to be rich with them. | ||
|- | |- | ||
| − | | T | + | | *T |
| Thr | | Thr | ||
| − | | [[Threonine]] | + | | [[Threonine]] |
| hydrophilic | | hydrophilic | ||
| 119.12 | | 119.12 | ||
| Line 286: | Line 286: | ||
| Behaves similarly to serine. | | Behaves similarly to serine. | ||
|- | |- | ||
| − | | V | + | | *V |
| Val | | Val | ||
| − | | [[Valine]] | + | | [[Valine]] |
| hydrophobic | | hydrophobic | ||
| 117.15 | | 117.15 | ||
| Line 297: | Line 297: | ||
| Behaves similarly to isoleucine and leucine. See isoleucine. | | Behaves similarly to isoleucine and leucine. See isoleucine. | ||
|- | |- | ||
| − | | W | + | | *W |
| Trp | | Trp | ||
| − | | [[Tryptophan]] | + | | [[Tryptophan]] |
| hydrophobic | | hydrophobic | ||
| 204.23 | | 204.23 | ||
Revision as of 15:44, 22 July 2006

An amino acid is an organic molecule with three main components: an amino group (-NH2), a carboxylic acid group (-COOH), and an R group (or side chain) unique to each amino acid.
Amino acids are the basic structural building blocks of proteins. Just as the letters of the alphabet can be combined in different ways to form an endless variety of words, a limited number of amino acids can be linked together in varying sequences to form a vast array of proteins. The unique three-dimensional shape of each protein, which results from the linear sequence of amino acids, determines the protein's specific function in the body.
Although over 100 amino acids exist in nature, our bodies require about 20 amino acids, called standard amino acids, for normal functioning. Eight of these standard amino acids are considered essential—that is, the body cannot synthesize them by itself and must obtain them from food.
Sources of amino acids
Standard amino acids
Twenty amino acids are known as standard amino acids or proteinogenic amino acids. As the name "proteinogenic" (literally, protein building) suggests, these amino acid are encoded by the standard genetic code and participate in the process of protein synthesis. They are formed from an mRNA template in a process called translation, by which genetic information, encoded in the form of nucleic acids, is translated into the amino acids essential for protein synthesis. Combinations of these amino acids produce every single protein required for the homeostasis (i.e., the maintenance of a stable internal environment) of the human body.
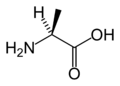
L-Alanine (Ala / A)
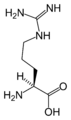
L-Arginine (Arg / R)
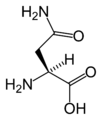
L-Asparagine (Asn / N)
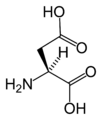
L-Aspartic acid (Asp / D)
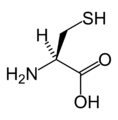
L-Cysteine (Cys / C)
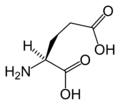
L-Glutamic Acid (Glu / E)
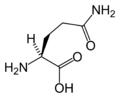
L-Glutamine (Gln / Q)
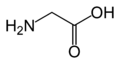
L-Glycine (Gly / G)
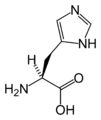
L-Histidine (His / H)
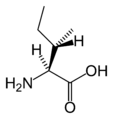
L-Isoleucine (Ile / I)
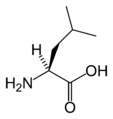
L-Leucine (Leu / L)
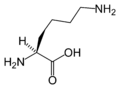
L-Lysine (Lys / K)
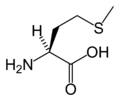
L-Methionine (Met / M)
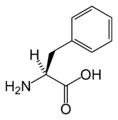
L-Phenylalanine (Phe / F)
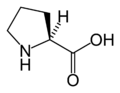
L-Proline (Pro / P)
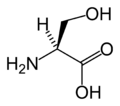
L-Serine (Ser / S)
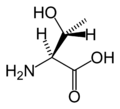
L-Threonine (Thr / T)
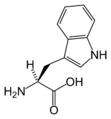
L-Tryptophan (Trp / W)
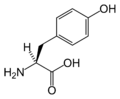
L-Tyrosine (Tyr / Y)
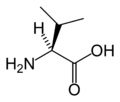
L-Valine (Val / V)

.png)


















Chemical structures of the 20 standard amino acids.
Essential amino acids
Eight of the 20 standard amino acids are called essential amino acids because they cannot be synthesized by the body from other compounds through chemical reactions; instead, they must be taken in with food. In humans, the essential amino acids are lysine, histidine, isoleucine, phenylalanine, leucine, methionine, tryptophan, threonine and valine. Arginine and histidine are generally considered essential only in children, whose undeveloped metabolisms are unable to synthesize them.
The remaining standard amino acids are nonessential; although they can be obtained from food, the body can also synthesize them as needed.
Nonstandard amino acids
Aside from the twenty standard amino acids and two special amino acids, selenocysteine and pyrrolysine, which are coded by DNA in an atypical manner, there are a vast number of nonstandard amino acids. These nonstandard or non-proteinogenic amino acids are either not found in proteins (like carnitine, GABA, or L-DOPA), or not coded for in the standard genetic code (like hydroxyproline and selenomethionine). The latter often result from posttranslational modification of proteins: the modification of standard amino acids after the protein has been formed in the translation stage of protein synthesis.
Some of these non-standard amino acids have been detected in meteorites, especially in a type known as carbonaceous chondrites, whose composition is considered to be representative of the solar nebula, or gaseous cloud, from which the solar system condensed. For example, over 79 amino acids were found in the primitive Murchison meteorite, a type of carbonaceous chondrite. Microorganisms and plants can also produce uncommon amino acids that can be found in peptidic antibiotics such as nisin, which is used as a food preservative.
The structure of amino acids
In biochemistry, the term "amino acid" is frequently used to refer specifically to alpha amino acids: those amino acids in which the amino and carboxylate groups are attached to the same carbon, the so-called α–carbon. The general structure of these proteinogenic alpha amino acids is:
R
|
H2N-C-COOH
|
H
where R represents a side chain specific to each amino acid. The exception to this basic structure is proline, whose side chain cyclizes onto the backbone, forming a ring structure in which a secondary amino group replaces the primary amino group.
When amino acids join together to form proteins, the basic amino group and acidic carboxyl group are neutralized. Instead, the side chains of the amino acids determine the acid-base properties of proteins. Thus, amino acids are usually classified by two main chemical properties of the side chain: the charge of the R group (which determines whether the protein acts like a weak acid or a weak base), and its polarity (or tendency to interact with water at a neutral pH).
These properties influence the amino acids' interaction with other structures within the protein as well as their interaction with other proteins. For example, soluble proteins have surfaces rich with polar amino acids like serine and threonine. In contrast, integral membrane proteins tend to have outer ring of hydrophobic amino acids (which do not tend to interact with water) that anchors them to the lipid bilayer. Similarly, proteins that have to bind to positively charged molecules have surfaces rich with negatively charged amino acids like glutamate and aspartate, while proteins binding to negatively charged molecules have surfaces rich with positively charged chains like lysine and arginine.
Isomers
Most amino acids occur in two possible optical isomers, called D and L. The L amino acids represent the vast majority of amino acids found in proteins. D amino acids are found in some proteins produced by exotic sea-dwelling organisms, such as cone snails. They are also abundant components of the proteoglycan cell walls of bacteria.
Amino acids function in protein synthesis

Two amino acids are linked together by a peptide bond, which forms when the basic amino group of one amino acid reacts with the acidic carboxyl group of a second amino acid. This condensation reaction (water loss) yields a peptide bond and a molecule of water. An amino acid residue is what is left of an amino acid once a molecule of water has been lost (an H+ from the amino group side and an OH- from the carboxylic side) in the formation of a peptide bond.
Proteins are then created by the polymerization of amino acids, a process in which amino acids are joined together in chains called, depending on their length, peptides or polypeptides.
Other biological roles of amino acids
In addition to their function in the formation of proteins, amino acids have other biologically-important roles. The standard amino acids glycine and glutamate, which are found in proteins, also function as neurotransmitters that respectively inhibit and amplify signals in the nervous system. The non-standard amino acid carnitine is used in lipid transport within the cell.
Many amino acids are used to synthesize molecules other than proteins:
- Tryptophan is a precursor of serotonin, a neurotransmitter important in the regulation of mood.
- Glycine is one of the reactants in the synthesis of porphyrins such as heme, which is a component of the hemoglobin molecules found in red blood cells.
- Arginine is used to synthesize the hormone nitric oxide.
Substances derived from amino acids have important uses in medicine and the food industry; for example:
- Aspartame (aspartyl-phenylalanine-1-methyl ester) is an artificial sweetener, and monosodium glutamate is a food additive that enhances flavor.
- 5-HTP (5-hydroxytryptophan) has been used to treat neurological problems associated with PKU (phenylketonuria), as well as depression (as an alternative to L-Tryptophan).
- L-DOPA (L-dihydroxyphenylalanine) is a drug used to treat the symptoms of Parkinson's disease.
Table of chemical properties
Following is a table listing the symbols and chemical properties of the standard amino acids. The mass listed is the weighted average of all common isotopes, and includes the mass of H2O. The one-letter symbol for an undetermined amino acid is X. The three-letter symbol Asx or one-letter symbol B means the amino acid is either asparagine or aspartic acid, whereas Glx or Z means either glutamic acid or glutamine. The three-letter symbol Sec or one-letter symbol U refers to selenocysteine. The letters J and O are not used.
Essential amino acids are marked with an asterisk.
| Abbrev. | Full Name | Side chain type | Mass | pI | pK1 (α-COOH) |
pK2 (α-+NH3) |
pKr (R) | Remarks | |
|---|---|---|---|---|---|---|---|---|---|
| A | Ala | Alanine | hydrophobic | 89.09 | 6.01 | 2.35 | 9.87 | Very abundant and versatile. It behaves fairly neutrally and can be located in both hydrophilic regions on the outside of the protein and hydrophobic interior areas. | |
| C | Cys | Cysteine | hydrophobic (Nagano, 1999) | 121.16 | 5.05 | 1.92 | 10.70 | 8.18 | The sulfur atom binds readily to heavy metal ions. Under oxidizing conditions, two cysteines can be joined together by a disulfide bond to form the amino acid cystine. When cystines are components of a protein, they enforce tertiary structure and makes the protein more resistant to unfolding and denaturation; disulphide bridges are therefore common in proteins that have to function in harsh environments, digestive enzymes (e.g., pepsin and chymotrypsin), structural proteins (e.g., keratin), and proteins too small to hold their shape on their own (eg. insulin). |
| D | Asp | Aspartic acid | acidic | 133.10 | 2.85 | 1.99 | 9.90 | 3.90 | Behaves similarly to glutamic acid. Carries a hydrophilic acidic group with strong negative charge. Usually is located on the outer surface of the protein, making it water-soluble. Binds to positively-charged molecules and ions, often used in enzymes to fix the metal ion. |
| E | Glu | Glutamic acid | acidic | 147.13 | 3.15 | 2.10 | 9.47 | 4.07 | Behaves similar to aspartic acid. Has longer, slightly more flexible side chain. |
| *F | Phe | Phenylalanine | hydrophobic | 165.19 | 5.49 | 2.20 | 9.31 | Phenylalanine, tyrosine, and tryptophan contain large rigid aromatic group on the side chain. These are the biggest amino acids. Like isoleucine, leucine and valine, they are hydrophobic and tend to orient towards the interior of the folded protein molecule. | |
| G | Gly | Glycine | hydrophobic | 75.07 | 6.06 | 2.35 | 9.78 | Because of the two hydrogen atoms at the α carbon, glycine is not optically active. It is the smallest amino acid, rotates easily, and adds flexibility to the protein chain. It is able to fit into the tightest spaces, e.g., the triple helix of collagen. | |
| H | His | Histidine | basic | 155.16 | 7.60 | 1.80 | 9.33 | 6.04 | In even slightly acidic conditions, protonation of the nitrogen occurs, changing the properties of histidine and the polypeptide as a whole. It is used by many proteins as a regulatory mechanism, changing the conformation and behavior of the polypeptide in acidic regions such as the late endosome or lysosome, enforcing conformation change in enzymes. |
| *I | Ile | Isoleucine | hydrophobic | 131.17 | 6.05 | 2.32 | 9.76 | Isoleucine, leucine and valine have large aliphatic hydrophobic side chains. Their molecules are rigid, and their mutual hydrophobic interactions are important for the correct folding of proteins, as these chains tend to be located inside of the protein molecule. | |
| *K | Lys | Lysine | basic | 146.19 | 9.60 | 2.16 | 9.06 | 10.54 | Behaves similarly to arginine. Contains a long flexible side-chain with a positively-charged end. The flexibility of the chain makes lysine and arginine suitable for binding to molecules with many negative charges on their surfaces. E.g., DNA-binding proteins have their active regions rich with arginine and lysine. The strong charge makes these two amino acids prone to be located on the outer hydrophilic surfaces of the proteins. |
| *L | Leu | Leucine | hydrophobic | 131.17 | 6.01 | 2.33 | 9.74 | Behaves similar to isoleucine and valine. See isoleucine. | |
| *M | Met | Methionine | hydrophobic | 149.21 | 5.74 | 2.13 | 9.28 | Always the first amino acid to be incorporated into a protein; sometimes removed after translation. Like cysteine, it contains sulfur, but with a methyl group instead of hydrogen. This methyl group can be activated, and is used in many reactions where a new carbon atom is being added to another molecule. | |
| N | Asn | Asparagine | hydrophilic | 132.12 | 5.41 | 2.14 | 8.72 | Neutralized version of aspartic acid. | |
| P | Pro | Proline | hydrophobic | 115.13 | 6.30 | 1.95 | 10.64 | Contains an unusual ring to the N-end amine group, which forces the CO-NH amide sequence into a fixed conformation. Can disrupt protein folding structures like α helix or β sheet, forcing the desired kink in the protein chain. Common in collagen, where it undergoes a posttranslational modification to hydroxyproline. Uncommon elsewhere. | |
| Q | Gln | Glutamine | hydrophilic | 146.15 | 5.65 | 2.17 | 9.13 | Neutralized version of glutamic acid. Used in proteins and as a storage for ammonia. | |
| R | Arg | Arginine | basic | 174.20 | 10.76 | 1.82 | 8.99 | 12.48 | Functionally similar to lysine. |
| S | Ser | Serine | hydrophilic | 105.09 | 5.68 | 2.19 | 9.21 | Serine and threonine have a short group ended with a hydroxyl group. Its hydrogen is easy to remove, so serine and threonine often act as hydrogen donors in enzymes. Both are very hydrophilic, therefore the outer regions of soluble proteins tend to be rich with them. | |
| *T | Thr | Threonine | hydrophilic | 119.12 | 5.60 | 2.09 | 9.10 | Behaves similarly to serine. | |
| *V | Val | Valine | hydrophobic | 117.15 | 6.00 | 2.39 | 9.74 | Behaves similarly to isoleucine and leucine. See isoleucine. | |
| *W | Trp | Tryptophan | hydrophobic | 204.23 | 5.89 | 2.46 | 9.41 | Behaves similarly to phenylalanine and tyrosine (see phenylalanine). Precursor of serotonin. | |
| Y | Tyr | Tyrosine | hydrophobic | 181.19 | 5.64 | 2.20 | 9.21 | 10.46 | Behaves similarly to phenylalanine and tryptophan (see phenylalanine). Precursor of melanin, epinephrine, and thyroid hormones. |
| Amino acid | Abbrev. | Side chain | Hydro- phobic | Polar | Charged | Small | Tiny | Aromatic or Aliphatic | van der Waals volume | Codon | Occurrence in proteins (%) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Alanine | Ala, A | -CH3 | X | - | - | X | X | - | 67 | GCU, GCC, GCA, GCG | 7.8 |
| Cysteine | Cys, C | -CH2SH | X | - | - | X | - | - | 86 | UGU, UGC | 1.9 |
| Aspartate | Asp, D | -CH2COOH | - | X | negative | X | - | - | 91 | GAU, GAC | 5.3 |
| Glutamate | Glu, E | -CH2CH2COOH | - | X | negative | - | - | - | 109 | GAA, GAG | 6.3 |
| Phenylalanine | Phe, F | -CH2C6H5 | X | - | - | - | - | Aromatic | 135 | UUU, UUC | 3.9 |
| Glycine | Gly, G | -H | X | - | - | X | X | - | 48 | GGU, GGC, GGA, GGG | 7.2 |
| Histidine | His, H | -CH2-C3H3N2 | - | X | positive | - | - | Aromatic | 118 | CAU, CAC | 2.3 |
| Isoleucine | Ile, I | -CH(CH3)CH2CH3 | X | - | - | - | - | Aliphatic | 124 | AUU, AUC, AUA | 5.3 |
| Lysine | Lys, K | -(CH2)4NH2 | - | X | positive | - | - | - | 135 | AAA, AAG | 5.9 |
| Leucine | Leu, L | -CH2CH(CH3)2 | X | - | - | - | - | Aliphatic | 124 | UUA, UUG, CUU, CUC, CUA, CUG | 9.1 |
| Methionine | Met, M | -CH2CH2SCH3 | X | - | - | - | - | - | 124 | AUG | 2.3 |
| Asparagine | Asn, N | -CH2CONH2 | - | X | - | X | - | - | 96 | AAU, AAC | 4.3 |
| Proline | Pro, P | -CH2CH2CH2- | X | - | - | X | - | - | 90 | CCU, CCC, CCA, CCG | 5.2 |
| Glutamine | Gln, Q | -CH2CH2CONH2 | - | X | - | - | - | - | 114 | CAA, CAG | 4.2 |
| Arginine | Arg, R | -(CH2)3NH-C(NH)NH2 | - | X | positive | - | - | - | 148 | CGU, CGC, CGA, CGG, AGA, AGG | 5.1 |
| Serine | Ser, S | -CH2OH | - | X | - | X | X | - | 73 | UCU, UCC, UCA, UCG, AGU,AGC | 6.8 |
| Threonine | Thr, T | -CH(OH)CH3 | X | X | - | X | - | - | 93 | ACU, ACC, ACA, ACG | 5.9 |
| Valine | Val, V | -CH(CH3)2 | X | - | - | X | - | Aliphatic | 105 | GUU, GUC, GUA, GUG | 6.6 |
| Tryptophan | Trp, W | -CH2C8H6N | X | - | - | - | - | Aromatic | 163 | UGG | 1.4 |
| Tyrosine | Tyr, Y | -CH2-C6H4OH | X | X | - | - | - | Aromatic | 141 | UAU, UAC | 3.2 |
Note: The pKa values of amino acids are typically slightly different when the amino acid is inside a protein. Protein pKa calculations are sometimes used to calculate the change in the pKa value of an amino acid in this situation
ReferencesISBN links support NWE through referral fees
- Doolittle, Russell F. 1989. Redundancies in protein sequences. In Predictions of Protein Structure and the Principles of Protein Conformation, ed. G.D. Fasman, 599-623. New York, NY: Plenum Press.
- Nelson, David L. and Michael M. Cox. 2000. Lehninger's Principles of Biochemistry, 3rd edition. New York, NY: Worth Publishers.
- Naganoa, Nozomi, Motonori Otaa and Ken Nishikawa. 1999. Strong hydrophobic nature of cysteine residues in proteins. FEBS Letters 458(1): 69-71. On the hydrophobic nature of cysteine.
External links
- Molecular Expressions: The Amino Acid Collection - Has detailed information and microscopy photographs of each amino acid.
- 22nd amino acid - Press release from Ohio State claiming discovery of a 22nd amino acid.
- Amino acid properties - Properties of the amino acids (a tool aimed mostly and molecular geneticists trying to understand the meaning of mutations)
- Synthesis of Amino Acids and Derivatives
- Right-handed amino acids were left behind
Credits
New World Encyclopedia writers and editors rewrote and completed the Wikipedia article in accordance with New World Encyclopedia standards. This article abides by terms of the Creative Commons CC-by-sa 3.0 License (CC-by-sa), which may be used and disseminated with proper attribution. Credit is due under the terms of this license that can reference both the New World Encyclopedia contributors and the selfless volunteer contributors of the Wikimedia Foundation. To cite this article click here for a list of acceptable citing formats.The history of earlier contributions by wikipedians is accessible to researchers here:
The history of this article since it was imported to New World Encyclopedia:
Note: Some restrictions may apply to use of individual images which are separately licensed.