Human evolution

Human evolution is that part of biological evolution concerning the emergence of humans as a distinct species. It is the subject of a broad scientific inquiry that seeks to understand and describe how this change and development occurred. The term "human," in the context of human evolution, refers to the genus Homo or more specifically the species Homo sapiens, but studies of human evolution usually include other hominins (humans, chimpanzees, and extinct ancestors), such as the australopithecines.
Human evolution theory deals with both the pattern of evolution (descent with modification, the non-causal relations between ancestral and descendant species) and the process of evolution (various theories involving mechanisms and causes for the pattern observed, including fundamental concepts such as natural selection, punctuated equilibrium, and design, and specific scenarios, such as those involving movement from trees, use of tools, "out of Africa," etc).
Substantial evidence has been marshaled for the fact that humans have descended from common ancestors by a process of branching (descent with modification) and for a primate origin of humans. However, proposals for the specific ancestral-descendant relationships and for the process leading to humans tend to be speculative. And, while the theory of natural selection typically is central to scientific explanations for the process, evidence for natural selection being the directive or creative force is limited to extrapolation from the microevolutionary level (changes within the level of species).
Historically, a major source of controversy has been the process by which humans have developed: whether by a non-directive process involving the creative force of natural selection acting on the chance occurrence of genetic variation (theory of natural selection); or by a directive process involving the creative force of a Creator God (and the non-random appearance of variation). Abrahamic religions believe in a single-point origin of modern humans, beginning with an Adam and Eve into whom God breathed life (added a spirit or soul to the material body). Whether the appearance of the human form was a chance event, brought about by natural selection and random variation, or a designed event brought about by a Creator God differs even among religious adherents.
This article will deal mainly with the pattern (descent with modification) of human evolution and not proposed mechanisms or causal explanations. The study of human evolution encompasses many scientific disciplines, most notably physical anthropology, linguistics, and genetics.
Terminology
- Hominoid: A hominoid is a member of the primate superfamily Hominoidea. The hominoids consist of the various species of gibbons (or "lesser apes"), as well as gorillas, chimpanzees, bonobos, orangutans, and humans (collectively referred to as the "great apes").
- Hominid: The term hominid technically refers to any member of the biological family Hominidae (the "great apes"), a group of primates that includes the extinct and extant humans, chimpanzees, gorillas, and orangutans. Originally, however, the term hominid was restricted to humans and their extinct relatives—those more closely related to humans than the other great apes, which were in a different family—and this definition of hominid is still used by many anthropologists and lay people.
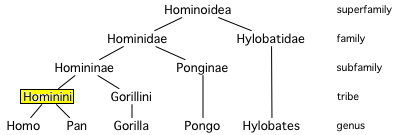 Extant Hominoid family tree
Extant Hominoid family tree - Hominin: A hominin is a member of the tribe Hominini: extinct and extant chimpanzees and humans.
- Hominan: A hominan is a member of the sub-tribe Hominina: humans and their extinct relatives.

Primate origin
There is substantial evidence for a primate origin of humans (Mayr 2001):
- Anatomical evidence: Human beings exhibit close anatomical similarities with the African apes, and particularly the chimpanzee. Compared to apes, the few unique physical characteristics of humans are the proportion of arms and legs, opposable thumbs, body hair, skin pigmentation, and size of the central nervous system, such as the forebrain.
- Fossil evidence: Numerous fossils have been found sharing human and primate characteristics.
- Molecular evidence: Human molecules are very similar to that of chimpanzees. In some, such as hemoglobin, they are virtually identical.
Indeed, the anatomical and biochemical similarity between chimpanzees and humans is so striking that some scientists have proposed that the two chimpanzee species, troglodytes and paniscus, belong with sapiens in the genus Homo, rather than in Pan. For example, comparisons between chimpanzees and humans in terms of protein sequences, allele differences, and DNA heteroduplex melting points show more than 98 percent identity (King and Wilson 1975; Wood 2006). Ebersberger et al. (2002) found a difference of only 1.24 percent when he aligned 1.9 million nucleotides of chimpanzee DNA and compared them with the corresponding human sequences in the human genome (Wood 2006). Other researchers found similar genetic similarities. Using a 4.97 million nucleotide portion of DNA from human chromosome 7 and comparing to chimpanzee orthologies yielded only 1.13 percent mismatches (Liu et al. 2003). A comparison of a rough draft of the chimpanzee genome with the human genome, for those sequences that could be aligned, averaged 1.23 percent nucleotide mismatches (The Chimpanzee Sequencing and Analysis Consortium 2005). Comparison of chimpanzee exons and human sequences yielded only 0.6 to 0.87 percent differences (Wildman et al. 2003; Nielsen et al. 2005).
Of course, the differences between apes and humans are likewise remarkable. Physiologically, chimpanzees rarely have heart attacks, are resistant to malaria caused by Plasmodium falciparum, and do not go through menopause (Wood 2006). Many cancers common to humans are not found in chimpanzees, and differences exist in terms of the genes for smell and also those that regulate the metabolism of amino acids. Behaviorally, human adult females, unlike any other species, are willing and capable of having sexual intercourse even during those times when they are not able to conceive and become pregnant. Beyond these physical measures, unlike chimpanzees, humans have complex languages (involving syntax and grammar), use symbols in communication, have developed complex technologies, practice and transmit religious belief, conduct scientific experiments, and so forth. Indeed, if one were to overlook the anatomical similarity, the gap between chimpanzees and humans in terms of culture, mental capacity, and various spiritual, emotional, and technological aspects is so large as to dwarf differences between chimpanzees and other animals. Theologically, humans are considered to be distinct from other animals.
Overview
The earliest delineated member of the genus Homo has generally been considered to be H. habilis, which lived from 2.33 to 1.44 million years ago, although some authorities do not consider it should be included in Homo, considering it more worthy, for example, to be retained in Australopithecus (Wood and Richmond 2000). There also are proposals for a new discovery, Homo gautengensis, to be considered the world's earliest recognized species of human, predating H. habilis (Viegas 2010).
Homo erectus is considered to have arrived around 1.8 million years ago, with fossils supporting its existence to 143,000 years ago. Often the early phase, from 1.8 to 1.25 mya, is considered either to be a separate species, H. ergaster, or it is seen as a subspecies of erectus, Homo erectus ergaster. That is, H. ergaster is viewed as possibly ancestral to H. erectus, or sharing a common ancestor with H. erectus, or is simply the African variety of H. erectus; regardless, it is widely considered to be the direct ancestor of later hominids such as Homo heidelbergensis, Homo sapiens, Neanderthals, Denisovans, and the Asian Homo erectus. Homo erectus and H. ergaster were the first of the hominina known to leave Africa. For example, H. erectus is known to have spread as far as Georgia, India, Sri Lanka, China and Java.
There is also support for the idea that that the numerous distinct species being recognized in the fossil record, such as H. erectus and H. habilis, are actually just morphological variation among members of a single evolving lineage among early members of the Homo genus, and that perhaps even only one species with a lot of variability emerged from Africa (Wilford 2013; Watson 2013; Lordkipanidze et al. 2013).
Modern human beings, Neanderthals, and Denisovans are believed to have shared a common ancestor about 400,000 to 500,000 years ago (Marshall 2013; Green et al. 2010). One theory is that these three groups all descended from Homo heidelbergenesis, which lived between 600,000 to 250,000 years ago (Marshall 2013) (other species suggested as ancestral are H. rhodesiensis and H. antecessor). One branch of H. heidelbergenesis are theorized to have left Africa about 400,000 years ago and split shortly thereafter to become Neanderthals, which settled in West Asia and Europe, and Denisovans, which settled farther to the east (NG 2013).
Neanderthals are considered to have lived from perhaps 400,000 years ago, with their appearance in the European fossil record variously estimated at 200,000 (Zimmer 2013) to 400,000 years ago (Green et al. 2010). Neanderthals disappeared from the fossil record about 30,000 years ago. Based on the DNA sequences for the nuclear genome of Neanderthals and modern humans, the population split between Neanderthals and modern humans took place 270,000 to 440,000 years ago (Reich et al. 2010).
Archaic Homo sapiens, the forerunner of anatomically modern humans, appeared between 400,000 and 250,000 years ago (O'Neil 2013). Anatomically modern humans are believed to have evolved from archaic Homo sapiens in the Middle Paleolithic, about 200,000 to 130,000 years ago (SA 2005; NG 2013), then migrated out of Africa some 50,000 to 100,000 years ago (Recent African Ancestory Theory) and replaced local populations of H. erectus, H. floresiensis, H. heidelbergenesis, and the Denisovan and Neanderthal populations.
The transition to behavioral modernity for Homo sapiens with the development of symbolic culture, language, and specialized lithic technology happened around 50,000 years ago according to many anthropologists (Mellars 2006), although some suggest a gradual change in behavior over a longer time span (Mcbrearty and Brooks 2000). Until about 50,000 to 40,000 years ago, the use of stone tools seems to have progressed stepwise: Each phase (habilis, ergaster, and neanderthal) started at a higher level than the previous one, but once that phase had started, further development was slow. After 50,000 years ago, in what Jared Diamond, author of The Third Chimpanzee, and other anthropologists characterize as a "Great Leap Forward," human culture apparently started to change at much greater speed: "Modern" humans started to bury their dead carefully, made clothing out of hides, developed sophisticated hunting techniques (such as pitfall traps, or driving animals to fall off cliffs), and made cave paintings. This speed-up of cultural change seems connected with the arrival of modern humans, Homo sapiens sapiens. Additionally, human culture began to become more technologically advanced, in that different populations of humans begin to create novelty in existing technologies. Artifacts such as fish hooks, buttons, and bone needles begin to show signs of variation among different population of humans, something what had not been seen in human cultures prior to 50,000 BP.
The Denisovans are not well delineated anatomically, given the very limited fossils found. The first fossils were discovered in 2008, when a small bone fragment of a finger was found. Two teeth were subsequently found. The lack of fossils has made anatomical representations of the group difficult. However, the DNA was preserved and was able to be extracted and has yielded excellent genetic analysis. As noted in a 2013 article in the Dartmouth Undergraduate Journal of Science, "Although Denisovans are thus far only represented by one finger bone and two teeth, they are currently the most well-known archaic human genetically – including Neanderthals of which there are hundreds of fossil records" (DUJS 2013). As a result, it was found that the Denisovans appear to be a unique group that shares a common origin with Neanderthals. DNA analysis further revealed that the Denisovans lived among and interbred with the ancestors of some present-day modern humans, with up to 6% of the DNA of Melanesians and Australian Aborigines deriving from Denisovans (Zimmer 2010; Callaway 2011).
In 2013, mitochondrial DNA was extracted from a 300,000- to 400,000-year-old sliver of hominin femur bone from Spain, which had been seen as either Neanderthal or Homo heidelbergensis. An almost complete mitochondrial genome was retrieved, the oldest human DNA sequenced. The DNA surprising yielded ancestral Denisonian DNA (Callaway 2013; Gibbons 2013).
Evidence and interpretation
There are two major scientific challenges in deducing the pattern of human evolution.
For one, the fossil record remains fragmentary. Mayr (2001) notes that no fossils of hominids have been found for the period between 6 and 13 million years ago (mya), the time when branching between the chimpanzee and human lineages is expected to have taken place. (While hominid refers to members of the "great ape" family, Hominidae, consisting of extinct and extant gorillas, chimpanzees, orangutans, and humans, Mayr appears to use it in the common anthropological content of animals more closely related to humans than other great apes, such as australopithecines.) Furthermore, as Mayr notes "most hominid fossils are extremely incomplete. They may consist of part of a mandible, or the upper part of a skull without face and teeth, or only part of the extremities." Even the famous "Lucy" finding (Australopithecus afarensis) was only a 40 percent complete female skeleton and lacked a head (Gould 1994).
Coupled with this is a recurrent problem that interpretation of fossil evidence is heavily influenced by personal beliefs and prejudices. Fossil evidence often allows a variety of interpretations, since the individual specimens may be reconstructed in a variety of ways (Wells 2000). As Mayr (2001) notes, "subjectivity is inevitable in the reconstruction of the missing parts," and virtually all hominid finds and interpretations are "somewhat controversial!" Wells (2000) recounts several examples where the pieces of fossils found offered a variety of reconstructions that were sometimes dramatically different, such as long face versus a short face, a heavy brow, a missing forehead. Different interpretations of two sections of a fossil skull and how to place one of those pieces led Roger Lewin to recount, "How you held it really depended on your preconceptions. It was very interesting what people did with it" (Wells 2000).
One significant development in elucidating the evolutionary relationships among species is the use of mitochondrial DNA (mtDNA) and nuclear DNA analysis. While for many fossils of ancient hominins the DNA cannot be recovered, in the case of more recent species, such as Denisova finds and Neanderthals, both mitochondrial and nuclear DNA has been recovered. With the sequences of modern humans also known, DNA analysis has been fruitful in providing significant insight into human origins and history.
History of paleoanthropology
The modern field of paleoanthropology (study of human origins) began in the nineteenth century with the discovery of "Neanderthal man" (the eponymous skeleton was found in 1856, but there had been finds elsewhere since 1830). However, fossils of Neanderthal's were widely misinterpreted as skeletons of modern humans with deformation or disease (Gould 1990).
The idea that humans are similar to certain great apes had been obvious to people for some time, but the idea of the biological evolution of species in general was not significantly advanced until after Charles Darwin published On the Origin of Species in 1859. Though Darwin's first book on evolution did not address the specific question of human evolution— "light will be thrown on the origin of man and his history" was all Darwin wrote on the subject— the implications of evolutionary theory were clear to contemporary readers. Debates between Thomas Huxley and Richard Owen focused on the idea of human evolution. Huxley convincingly illustrated many of the similarities and differences between humans and apes in his 1863 book Evidence as to Man's Place in Nature. By the time Darwin published his own book on the subject, Descent of Man, it was already a well-known interpretation of his theory—and the interpretation helped make the theory of natural selection highly controversial. Even many of Darwin's original supporters (such as Alfred Russel Wallace and Charles Lyell) balked at the idea that human beings could have evolved their apparently boundless mental capacities and moral sensibilities through natural selection.
Since the time of Carolus Linnaeus, the great apes were considered the closest relatives of human beings, based on morphological similarity. In the nineteenth century, it was speculated that their closest living relatives were chimpanzees and gorillas, and based on the natural range of these creatures, it was surmised humans share a common ancestor with other African apes and that fossils of these ancestors would ultimately be found in Africa.
It was not until the 1920s that hominin fossils were discovered in Africa. In 1924, Raymond Dart described Australopithecus africanus. The type specimen was the Taung Child, an australopithecine infant discovered in a cave deposit being mined for concrete at Taung, South Africa. The remains were a remarkably well-preserved tiny skull and an endocranial cast of the individual's brain. Although the brain was small (410 cm3), its shape was rounded, unlike that of chimpanzees and gorillas, and more like a modern human brain. Also, the specimen exhibited short canine teeth, and the position of the foramen magnum (the hole in the skull where the spine enters) was evidence of bipedal locomotion. All of these traits convinced Dart that the Taung baby was a bipedal human ancestor, a transitional form between apes and humans. Another 20 years would pass before Dart's claims were taken seriously, following the discovery of more fossils that resembled his find. The prevailing view of the time was that a large brain evolved before bipedality. It was thought that intelligence on par with modern humans was a prerequisite to bipedalism.
The australopithecines, which now are widely seen as direct ancestors leading to the human lineage, have been classified as either gracile (Australopithecus genus) or robust (Paranthropus genus). However, some consider the robust species aethiopicus, boisei, and robustus as included within the genus Australopithecus. In the 1930s, when the robust specimens were first described, the Paranthropus genus was used. During the 1960s, the robust variety was moved into Australopithecus. The recent trend has been back to the original classification as a separate genus. Human evolution/Species chart
Before Homo
Evolutionary history of the primates can be traced back 65 million years. The oldest known primate-like mammal species, the Plesiadapis, came from North America, but they were widespread in Eurasia and Africa during the tropical conditions of the Paleocene and Eocene.
With the beginning of modern climates, marked by the formation of the first Antarctic ice in the early Oligocene around 40 million years ago, primates went extinct everywhere but Africa and southern Asia.
| Paleogene period | ||
|---|---|---|
| Paleocene epoch | Eocene epoch | Oligocene epoch |
| Danian | Selandian Thanetian |
Ypresian | Lutetian Bartonian | Priabonian |
Rupelian | Chattian |
Fossil evidence found in Germany 20 years ago (Kordon and Begun 2001, Heizmann and Begun 2001) was determined to be about 16.5 million years old, some 1.5 million years older than similar species from East Africa. It suggests that the great ape and human lineage first appeared in Eurasia and not Africa. The discoveries suggest that the early ancestors of the hominids (the family of great apes and humans) migrated to Eurasia from Africa about 17 million years ago, just before these two continents were cut off from each other by an expansion of the Mediterranean Sea.
Heizmann and Begun (2001) hold that the great apes flourished in Eurasia and that their lineage leading to the African apes and humans—Dryopithecus—migrated south from Europe or Western Asia into Africa. The surviving tropical population, which is seen most completely in the upper Eocene and lowermost Oligocene fossil beds of the Fayum depression southwest of Cairo, is held to have given rise to all living primates—lemurs of Madagascar, lorises of Southeast Asia, galagos or "bush babies" of Africa, and the anthropoids: platyrrhines or New World monkeys, and catarrhines or Old World monkeys and the great apes and humans.
The earliest known catarrhine, or Old World monkey, is Kamoyapithecus from uppermost Oligocene at Eragaleit in the northern Kenya rift valley, dated to 24 mya (millions years ago). Its ancestry is generally thought to be close to such genera as Aegyptopithecus, Propliopithecus, and Parapithecus from the Fayum, at around 35 mya. There are no fossils from the intervening 11 million years.
No near ancestor to South American platyrrhines, whose fossil record begins at around 30 Ma, can be identified among the North African fossil species, and possibly lies in other forms that lived in West Africa that were caught up in the still-mysterious transatlantic sweepstakes that sent primates, rodents, boa constrictors, and cichlid fishes from Africa to South America sometime in the Oligocene.
In the early Miocene, after 22 mya, many kinds of arboreally adapted primitive catarrhines from East Africa appeared. Because the fossils at 20 mya include fragments attributed to Victoriapithecus, the earliest cercopithecoid, the other forms are (by default) grouped as hominoids, without clear evidence as to which are closest to living apes and humans. Among the presently recognized genera in this group, which ranges up to 13 mya, one finds Proconsul, Rangwapithecus, Dendropithecus, Limnopithecus, Nacholapithecus, Equatorius, Nyanzapithecus, Afropithecus, Heliopithecus, and Kenyapithecus, all from East Africa. The presence of other generalized non-cercopithecids of middle Miocene age from sites far distant—Otavipithecus from cave deposits in Namibia, and Pieroloapithecus and Dryopithecus from France, Spain, and Austria—is evidence of a wide diversity of forms across Africa and the Mediterranean basin during the relatively warm and equable climatic regimes of the early and middle Miocene. The youngest of the Miocene hominoids, Oreopithecus, is from 9 mya coal beds in Italy.
| Tertiary sub-era | Quaternary sub-era | |||
|---|---|---|---|---|
| Neogene period | ||||
| Miocene | Pliocene | Pleistocene | Holocene | |
| Aquitanian | Burdigalian | Zanclean | Early | |
| Langhian | Serravallian | Piacenzian | Middle | |
| Tortonian | Messinian | Gelasian | Late | |
Gibbons and orangutans. Molecular evidence indicates that the lineage of gibbons (family Hylobatidae) became distinct between 18 and 12 mya, and that of orangutans (subfamily Ponginae) at about 12 mya. We have no fossils that clearly document the ancestry of gibbons, which may have originated in a so far unknown South East Asian hominoid population, but fossil proto-orangutans may be represented by Ramapithecus from India and Griphopithecus from Turkey, dated to around 10 mya.
Gorillas, chimpanzees, and hominans. Molecular evidence further suggests that between 8 and 4 mya, first the gorillas, and then the chimpanzee (genus Pan) split off from the line leading to the modern humans. However, there is no fossil record of either group of African great apes. One explanation is that bones do not fossilize well in rainforest environments. Patterson et al. (2006) reported on genetic evidence that humans and chimpanzees probably diverged less than 6.3 mya. More controversially, they speculated on the basis of the evidence that after divergence of the lineages, interbreeding again took place, before the final split about 5.3 mya.
Hominines, however, seem to have been one of the mammal groups (as well as antelopes, hyenas, dogs, pigs, elephants, and horses) that adapted to the open grasslands as soon as this biome appeared, due to increasingly seasonal climates, about 8 mya, and their fossils are relatively well known. The earliest are Sahelanthropus tchadensis (7-6 mya) and Orrorin tugenensis (6 mya), followed by:
- Ardipithecus (5.5-4.4 mya), with species Ar. kadabba and Ar. ramidus;
- Australopithecus (4-2 mya), with species Au. anamensis, Au. afarensis, Au. africanus, Au. bahrelghazali, and Au. garhi;
- Paranthropus (3-1.2 mya), with species P. aethiopicus, P. boisei, and P. robustus;
- Homo (2 mya-present).
Genus Australopithecus
The Australopithecus genus evolved in eastern Africa around 4 million years ago before spreading throughout the continent and eventually becoming extinct 2 million years ago. During this time period various forms of australopiths existed, including Australopithecus anamensis, A. afarensis, A. sediba, and A. africanus. There is still some debate amongst academics whether certain African hominid species of this time, such as A. robustus and A. boisei, constitute members of the same genus; if so, they would be considered to be robust australopiths while the others would be considered gracile australopiths. However, if these species do indeed constitute their own genus, then they may be given their own name, the Paranthropus.
- Australopithecus (4–1.8 Ma), with species Au. anamensis, Au. afarensis, Au. africanus, Au. bahrelghazali, Au. garhi, and Au. sediba;
- Kenyanthropus (3–2.7 Ma), with species Kenyanthropus platyops;
- Paranthropus (3–1.2 Ma), with species P. aethiopicus, P. boisei, and P. robustus;
Australopithecus afarensis and Australopithecus africanus are among the most famous of the extinct hominids. The australopithecines are now thought to be immediate ancestors of the genus Homo, the group to which modern humans belong. Both australopithecines and Homo sapiens are part of the tribe Hominini.
The genus Homo

In modern taxonomy, Homo sapiens is the only extant (living) species of its genus, Homo. However, the ongoing study of the origins of Homo sapiens often demonstrates that there were other Homo species, all of which are now extinct. While some of these other species might have been ancestors of H. sapiens, many were likely "cousins," having speciated away from our ancestral line. There is not yet a consensus as to which of these groups should count as separate species and which as subspecies of another species. In some cases, this is due to the paucity of fossils; in other cases, it is due to the slight differences used to classify species in the Homo genus.
The word homo is Latin for "person," chosen originally by Carolus Linnaeus in his classification system. It is often translated as "man," although this can lead to confusion, given that the English word "man" can be generic like homo, but can also specifically refer to males. Latin for "man" in the gender-specific sense is vir, cognate with "virile" and "werewolf." The word "human" is from humanus, the adjectival form of homo.
Homo habilis and H. gautengensis
H. habilis lived from about 2.4 to 1.5 million years ago (mya). H. habilis, the oldest known species of the genus Homo, appeared in South and East Africa in the late Pliocene or early Pleistocene, 2.5–2 mya, when it is considered to have diverged from the Australopithecines. H. habilis had smaller molars and larger brains than the Australopithecines, and made tools from stone and perhaps animal bones. One of the first known hominids, it was nicknamed "handy man" by its discoverer, Louis Leakey. Some scientists have proposed moving this species out of Homo and into Australopithecus due to the morphology of its skeleton being more adapted to living on trees rather than to moving on two legs like Homo sapiens (Wood and Collard 1999).
It was considered to be the first species of the genus Homo until May 2010, when a new species, Homo gautengensis was discovered in South Africa, that most likely arose earlier than Homo habilis (Viegas 2010).
Homo rudolfensis and Homo georgicus
These are proposed species names for fossils from about 1.9-1.6 mya, the relation of which with H. habilis is not yet clear.
- H. rudolfensis refers to a single, incomplete skull from Kenya. Some consider this another H. habilis.
- H.georgicus, from the country of Georgia, may be an intermediate form between H. habilis and H. erectus or a subspecies of H. erectus.
Homo ergaster and Homo erectus
The first fossils of Homo erectus were discovered by Dutch physician Eugene Dubois in 1891 on the Indonesian island of Java. He originally gave the material the name Pithecanthropus erectus based on its morphology, which he considered to be intermediate between that of humans and apes.
H. erectus lived from about 1.8 mya to 70,000 years ago. Homo erectus was apparently very successful, with fossils found in Africa, Asia (Indonesia, and China), Georgia (Caucasus region of Europe), and eastern and southern Africa (Mayr 2001). A famous example of Homo erectus is Peking Man.
Often the early phase, from 1.8 to 1.25 mya, is considered either to be a separate species, H. ergaster, or it is seen as a subspecies of erectus, Homo erectus ergaster. That is, the differences between the early populations of H. erectus, found in Africa, and the later populations, found in Asia, Europe, and Africa, are substantial enough for the separation by many researchers into two different groups: the early African H. ergaster or H. erectus ergaster, and the mainly Asian populations H. erectus (Smithsonian 2007b). Many paleoanthropologists now use the term Homo ergaster only for the non-Asian forms of this group, and reserve Homo erectus only for those fossils that are found in Asia and meet certain skeletal and dental requirements, which differ slightly from H. ergaster.
Homo erectus and H. ergaster were the first of the hominina known to leave Africa.
In the Early Pleistocene, 1.5–1 mya, in Africa, Asia, and Europe, presumably, Homo habilis evolved larger brains and made more elaborate stone tools; these differences and others are sufficient for anthropologists to classify them as a new species, H. erectus. In addition H. erectus was the first human ancestor to walk truly upright. This was made possible by the development of locking knees and a different location of the foramen magnum (the hole in the skull where the spine enters). They may have used fire to cook their meat.
Homo cepranensis and Homo antecessor
These are proposed as species that may be intermediate between H. erectus and H. heidelbergensis.
- H. cepranensis refers to a single skull cap from Italy, estimated to be about 800,000 years old.
- H. antecessor is known from fossils from Spain and England that are 800,000-500,000 years old.
Homo heidelbergensis
H. heidelbergensis (Heidelberg Man) lived from about 800,000 to about 300,000 years ago. Also proposed as Homo sapiens heidelbergensis or Homo sapiens paleohungaricus.
Homo rhodesiensis, and the Gawis cranium
- H. rhodesiensis, estimated to be 300,000-125,000 years old, most current experts believe Rhodesian Man to be within the group of Homo heidelbergensis though other designations such as Archaic Homo sapiens and Homo sapiens rhodesiensis have also been proposed.
- In February 2006, a fossil, the Gawis cranium, was found that might possibly be a species intermediate between H. erectus and H. sapiens or one of many evolutionary dead ends. The skull from Gawis, Ethiopia, is believed to be 500,000-250,000 years old. Only summary details are known, and no peer reviewed studies have been released by the finding team. Gawis man's facial features suggest it's either an intermediate species or an example of a "Bodo man" female (IU 2006).
Neanderthals
H. neanderthalensis, alternatively designated as Homo sapiens neanderthalensis (Harvati 2003), lived in Europe and Asia from 400,000 to about 30,000 years ago. Evidence from sequencing mitochondrial DNA indicated that no significant gene flow occurred between H. neanderthalensis and H. sapiens, and, therefore, the two were separate species that shared a common ancestor about 660,000 years ago (Krigs et al. 1997; Green et al. 2008; Serre et al. 2004). However, the 2010 sequencing of the Neanderthal genome indicated that Neanderthals did indeed interbreed with anatomically modern humans circa 45,000 to 80,000 years ago (at the approximate time that modern humans migrated out from Africa, but before they dispersed into Europe, Asia and elsewhere) (Green et al. 2010).
Nearly all modern non-African humans have 1% to 4% of their DNA derived from Neanderthal DNA (Green et al. 2010), and this finding is consistent with recent studies indicating that the divergence of some human alleles dates to one Ma, although the interpretation of these studies has been questioned (Gutiérrez et al. 2002; Hebsgaard et al. 2007). Competition from Homo sapiens probably contributed to Neanderthal extinction (Banks et al. 2008; Hortola and Martinez-Navarro 2012). They could have co-existed in Europe for as long as 10,000 years, during which human populations exploded, vastly outnumbering Neanderthals, possibly outcompeting them by sheer numerical strength (Mellars and French 2011).
Denisovans
Denisovans are an extinct hominid group more closely related to the Neanderthals than modern humans and identified from the nuclear and mitochondrial DNA sequences of the roughly 50,000-year-old (or older) fossils found in Siberia. The fossils unearthed from the Denisova Cave in the Altai Mountains of southern Siberia are quite limited in number: a pinkie bone (distal manual phalanx) and two teeth (molars) from three different Denisovan individuals, as of 2013. However, the entire genome has been sequenced and this DNA sequence shows the Denisovans to be a distinct group that shares a common ancestor with Neanderthals. It is believed they were more prevalent in East Asia while Neanderthals dominated Europe and western Asia.
The issue of whether the Denisovans are a unique species or a subspecies of Homo sapiens (Homo sapiens ssp. 'Denisova) has not been settled, just as it has not been settled for the Neanderthals.
Homo floresiensis?
H. floresiensis is the name given to a proposed species that lived about 100,000-12,000 years ago (announced October 28, 2004 in the science journal Nature) (Brown et al. 2004). It is based on fossil findings, with the main find a fossil believed to be a woman about 30 years of age. Found in 2003, it has been dated to approximately 18,000 years old. The living woman was estimated to be one meter in height, with a brain volume of just 380 cm3 (which can be considered small even for a chimpanzee). The species has been nicknamed "hobbit" for its small size, possibly a result of insular (island) dwarfism (Brown et al. 2004).
H. floresiensis is intriguing both for its size and its age, being a concrete example of what some consider a recent species of the genus Homo that exhibits derived traits not shared with modern humans. In other words, H. floresiensis is postulated to share a common ancestor with modern humans, but split from the modern human lineage and followed a distinct evolutionary path.
However, there is an ongoing debate over whether H. floresiensis is indeed a separate species (Argue et al. 2006). Some scientists presently believe that H. floresiensis was a modern H. sapiens suffering from pathological dwarfism (Martin et al. 2006). This hypothesis is supported in part, because the modern humans who live on Flores, the island where the fossil was found, are pygmies. This coupled with pathological dwarfism could indeed create a hobbit-like human. The other major attack on H. floresiensis as a separate species is that it was found with tools only associated with H. sapiens.
The hypothesis of pathological dwarfism, however, fails to explain additional anatomical features that are unlike those of modern humans (diseased or not) but much like those of ancient members of our genus. Aside from cranial features, these features include the form of bones in the wrist, forearm, shoulder, knees, and feet. Additionally, this hypothesis fails to explain the find of multiple examples of individuals with these same characteristics, indicating they were common to a large population, and not limited to one individual.
Homo sapiens
H. sapiens ("sapiens" means wise or intelligent) have lived from about 250,000 years ago to the present. Between 400,000 years ago and the second interglacial period in the Middle Pleistocene, around 250,000 years ago, the trend in cranial expansion and the elaboration of stone tool technologies developed, providing evidence for a transition from H. erectus to H. sapiens.
The direct evidence suggests there was a migration of H. erectus out of Africa, then a further speciation of H. sapiens from H. erectus/H. ergaster in Africa. Then a subsequent migration within and "out of Africa" eventually replaced the earlier dispersed H. erectus. This migration and origin theory is usually referred to as the recent single-origin theory or "Out of Africa Hypothesis". However, the current evidence does not preclude multiregional evolution or some mixtrue of the migrant H. sapiens with existing Homo populations. This is a hotly debated topic in paleoanthropology.
Based on molecular evidence, the calculation of the time of divergence of all modern human populations from a common ancestor typically yields dates around 200,000 years (Smithsonian 2007a). The oldest fossil evidence for modern humans is 130,000 years old in Africa and sometime before 90,000 years old in the Near East (Smithsonian 2007a). Some consider H. sapiens idaltu from Ethiopia, an extinct subspecies which lived from about 160,000 years ago, to be the oldest known anatomically modern human.
Notably, however, about 50,000 to 40,000 years ago, human beings appeared to have taken a Great Leap Forward, when human culture apparently changed at a much greater speed. It seems connected to the arrival of modern humans beings: Homo sapiens sapiens. (See modern man and the great leap forward.)
The Cro-Magnons form the earliest known European examples of Homo sapiens sapiens. The term falls outside the usual naming conventions for early humans and is used in a general sense to describe the oldest modern people in Europe. Cro-Magnons lived from about 40,000 to 10,000 years ago in the Upper Paleolithic period of the Pleistocene epoch. For all intents and purposes these people were anatomically modern, only differing from their modern day descendants in Europe by their slightly more robust physiology and larger brain capacity than that of modern humans. When they arrived in Europe about 40,000 years ago, they brought with them sculpture, engraving, painting, body ornamentation, music, and the painstaking decoration of utilitarian objects.
Current research establishes that human beings are highly genetically homogeneous, meaning that the DNA of individual Homo sapiens is more alike than usual for most species. Distinctive genetic characteristics have arisen, however, primarily as the result of small groups of people moving into new environmental circumstances. Such small groups are initially highly inbred, allowing the relatively rapid transmission of traits favorable to the new environment. These adapted traits are a very small component of the Homo sapiens genome and include such outward "racial" characteristics as skin color and nose form in addition to internal characteristics such as the ability to breathe more efficiently in high altitudes.
Abrahamic religions believe in a single-point origin of modern humans, beginning with an original couple, Adam and Eve, into whom God breathed life (added a spirit or soul to the material body).
Archaic human admixture
Over the course of human history, archaic human admixture has occurred with modern humans (Homo sapiens), namely through interbreeding of modern humans with Neanderthals and Denisovans respectively.
A detailed comparison of the Denisovan, Neanderthal, and human genomes has revealed evidence for a complex web of interbreeding among the lineages. Through such interbreeding, 17% of the Denisova genome represents DNA from the local Neanderthal population, while evidence was also found of a contribution to the nuclear genome from an ancient hominin lineage yet to be identified (Pennisi 2013). An estimated 1% to 4% of the DNA in Eurasians is non-modern and shared with Neanderthals rather than with sub-Saharan Africans. In Oceanian and some Southeast Asian populations, Denisovan admixture has also been observed. An estimated 4 to 6% of Melanesian DNA is derived from Denisovans.
Various theories of Neanderthal admixture in modern human DNA—i.e. the result of interbreeding of Neanderthals and anatomically modern humans during the Middle Paleolithic—have been debated throughout the 20th century, and in terms of genetics throughout the 2000s (Fagundes et al. 2007; Hodgson and Disotell 2008)). A 2010 draft sequence publication on the Neanderthal genome indicates that Neanderthals share genetic lineages with Eurasian populations and not with Sub-Saharan African populations (Green et al. 2010). According to the study, this scenario is best explained by gene flow from Neanderthals to modern humans after humans emerged from Africa and possibly before the divergence of the Eurasian groups (Green et al. 2010). The data show that 1–4% of the DNA in Europeans and Asians (e.g. French, Han Chinese, and Papua New Guinean) is non-modern and shared with Neanderthals rather than with Sub-Saharan Africans (e.g. Yoruba and San) (Green et al. 2010). Though less parsimonious than gene flow, early ancestors of Eurasians may have been more closely related to Neanderthals than those of Africans to Neanderthals, a result from incomplete genetic homogenization due ancient population sub-structure in Africa when Neanderthals diverged (Green et al. 2010). No evidence of Neanderthal mitochondrial DNA have been found in modern humans (Krings et al. 1997; Serre et al. 2004; Wall and Hammer 2006). This would suggest that successful Neanderthal admixture happened paternally rather than maternally (Mason and Short 2011; Wang et al. 2013). Possible hypotheses are that Neanderthal mtDNA had detrimental mutations that led to the extinction of carriers, that the hybrid offspring of Neanderthal mothers were raised in Neanderthal groups and became extinct with them, or that female Neanderthals and male modern humans did not produce fertile offspring (Mason and Short 2011).
Analysis of genomes of modern humans show that they mated with at least two groups of ancient humans: Neanderthals (more similar to those found in the Caucasus than those from the Altai region) (Pennisi 2013) and Denisovans )Green et al. 2010). Tests comparing the Denisova hominin genome with those of six modern humans—a ǃKung from South Africa, a Nigerian, a Frenchman, a Papua New Guinean, a Bougainville Islander, and a Han Chinese—showed that between 4% and 6% of the genome of Melanesians (represented by the Papua New Guinean and Bougainville Islander) derives from a Denisovan population. This DNA was possibly introduced during the early migration to Melanesia. These findings are in concordance with the results of other comparison tests that show a relative increase in allele sharing between the Denisovan and the Aboriginal Australian genome, compared to other Eurasians and African populations; however it has been observed that Papuans, the population of Papua New Guinea, have more allele sharing than Aboriginal Australians (Rasmussen et al. 2011).
Mitochondrial Eve and Y-chromosomal Adam
Geneticists have identified individuals dubbed "Mitochondrial Eve" and "Y-chromosomal Adam," with Mitochondrial Eve the common matrilineal ancestor of all humans alive today and Y-chromosomal Adam the common patrilineal ancestor who lived many millennia after Mitochondrial Eve. (That a common patrilineal ancestor should have lived significantly after a matrilineal one is in concordance with Bible history, which gives Noah as a more recent common patrilineal ancestor than Adam.) In the founding work of this area of investigation, Cann et al. (1987) compared mitochondrial DNA sampled from women whose ancestors came from different part of the world. and concluded that Africa was the most likely root of human ancestry and that human ancestors left Africa between between 180,000 and 230,000 years ago.
Note that Mitochondrial Eve is the most recent common matrilineal ancestor, and Y-chromosomal Adam the most common patrilineal ancestor, not the original man and woman, and not the most recent common ancestor (MRCA) of all humans. All living humans can trace their ancestry back to the MRCA via at least one of their parents. That is, the MRCA's offspring have led to all living humans via sons and daughters. However, Mitochondrial Eve must be traced only through female lineages, so she is estimated to have lived much longer ago than the MRCA. While Mitochondrial Eve is thought to have been living around 140,000 years ago (Dawkins 2004) or between 100,000 and 200,000 years ago (Rhode 2005), and Y-chromosomal Adam about 35,000 to 89,000 years ago (Rhode 2005), the MRCA is estimated to have been living only around 2,000 to 5,000 years ago (Rohde et al. 2004, Rhode 2005).
Starting with "the" MRCA at around 3,000 years ago, one can trace all ancestors of the MRCA backward in time. At every ancestral generation, more and more ancestors (via both paternal and maternal lines) of MRCA are found. These ancestors are by definition also common ancestors of all living people. Eventually, there will be a point in the past where all humans can be divided into two groups: Those who left no descendants today and those who are common ancestors of all living humans today. This point in time is termed the "identical ancestors point" and is estimated to be between 5,000 and 15,000 years ago. Since Mitochondrial Eve is estimated to have lived more than hundred thousand years before the identical ancestors point, every contemporary woman of hers is either not an ancestor of all living people, or a common ancestor of all living people (Dawkins 2004; Rohde 2005).
Comparative table of Homo species
- Bolded species names indicate the existence of numerous fossil records.
| species | lived when (MYA) | lived where | adult length (m) | adult weight (kg) | brain volume (cm³) | fossil record | discovery / publication of name |
|---|---|---|---|---|---|---|---|
| H. habilis | 2.5–1.5 | Africa | 1.0–1.5 | 30–55 | 600 | many | 1960/1964 |
| H. rudolfensis | 1.9 | Kenya | 1 skull | 1972/1986 | |||
| H. georgicus | 1.8–1.6 | Georgia | 600 | few | 1999/2002 | ||
| H. ergaster | 1.9–1.25 | E. and S. Africa | 1.9 | 700–850 | many | 1975 | |
| H. erectus | 2(1.25)–0.3 | Africa, Eurasia (Java, China, Caucasus) | 1.8 | 60 | 900–1100 | many | 1891/1892 |
| H. cepranensis | 0.8? | Italy | 1 skull cap | 1994/2003 | |||
| H. antecessor | 0.8–0.35 | Spain, England | 1.75 | 90 | 1000 | 3 sites | 1997 |
| H. heidelbergensis | 0.6–0.25 | Europe, Africa, China | 1.8 | 60 | 1100–1400 | many | 1908 |
| H. neanderthalensis | 0.23–0.03 | Europe, W. Asia | 1.6 | 55–70 (heavily built) | 1200-1700 | many | (1829)/1864 |
| H. rhodesiensis | 0.3–0.12 | Zambia | 1300 | very few | 1921 | ||
| H. sapiens sapiens | 0.25–present | worldwide | 1.4–1.9 | 55–80 | 1000–1850 | still living | —/1758 |
| H. sapiens idaltu | 0.16 | Ethiopia | 1450 | 3 craniums | 1997/2003 | ||
| H. floresiensis? | 0.10–0.012 | Indonesia | 1.0 | 25 | 400 | 7 individuals | 2003/2004 |
Out of Africa and multiregional positions
There are two dominant, and one might say polarizing, general views on the issue of human origins, the Out of Africa position and the multiregional position.
The Out of Africa, or Out of Africa II, or replacement model holds that after there was a migration of Homo erectus (or H. ergaster) out of Africa and into Europe and Asia, these populations did not subsequently contribute significant amounts of genetic material (or, some say, contributed absolutely nothing) to later populations along the lineage to Homo sapiens (Kreger 2005). Later, approximately 200,000 years ago, there was a second migration of hominids out of Africa, and this was modern H. sapiens that replaced the populations that then occupied Europe and Asia (Kreger 2005). This view maintains a specific speciation event that led to H. sapiens in Africa, and this is the modern human.
The multiregional or continuity camp hold that since the origin of H. erectus, there have been populations of hominids living in the Old World and that these all contributed to successive generations in their regions (Kreger 2005). According to this view, hominids in China and Indonesia are the most direct ancestors of modern East Asians, those in Africa are the most direct ancestors of modern Africans, and the European populations either gave rise to modern Europeans or contributed significant genetic material to them, while their origins were in Africa or West Asia (Kreger 2005). There is genetic flow to allow for the maintenance of one species, but not enough to prevent racial differentiation.
There are various combinations of these ideas. Overall, the disagreement between these two camps has caused "fairly severe strife within the paleoanthropological community" (Kreger 2005). Multiregionalism is often "portrayed as a racist theory," while Out of Africa II "has often been portrayed as a religiously motivated idea" that strives to align with the biblical story of Genesis (Kreger 2005).
Use of tools
Using tools is a sign of intelligence. Some scientists speculate that tool use also played a role in the origin of modern humans. Over the past 2 or 3 million years, human brain size has increased threefold. A brain needs a lot of energy: The brain of a modern human uses about 20 Watts (about 400 calories per day), one fifth of total human energy consumption. Early hominoids, like apes, were considered to have been essentially plant eaters (fruit, leaves, roots), their diet only occasionally supplemented by meat (often from scavenging). However, plant food in general yields considerably less energy and nutritive value than meat. Therefore, being able to hunt for large animals, which was only possible by using tools such as spears, would have made it possible for humans to sustain larger and more complex brains, which in turn allowed them to develop yet more intelligent and efficient tools.
Precisely when early humans started to use tools is difficult to determine, because the more primitive these tools were (for example, sharp-edged stones), the more difficult it is to decide whether they are natural objects or human artifacts. There is some evidence that the australopithecines (4 mya) may have used broken bones as tools, but this is debated.
Stone tools
Stone tools are first attested around 2.6 mya, when H. habilis in Eastern Africa used so-called pebble tools: Choppers made out of round pebbles that had been split by simple strikes. This marks the beginning of the Paleolithic, or Old Stone Age; its end is argued to be the end of the last Ice Age, around 10,000 years ago. The Paleolithic is subdivided into the Lower Paleolithic (Early Stone Age, ending around 350,000–300,000 years ago), the Middle Paleolithic (Middle Stone Age, until 50,000–30,000 years ago), and the Upper Paleolithic.
The period from 700,000–300,000 years ago is also known as the Acheulean, when H. ergaster (or erectus) made large stone hand-axes out of flint and quartzite, at first quite rough (Early Acheulian), later "retouched" by additional, more subtle strikes at the sides of the flakes.
After 350,000 BP (Before Present), a more refined, the so-called Levallois, technique was developed. It consisted of a series of consecutive strikes, by which scrapers, slicers ("racloirs"), needles, and flattened needles were made. Finally, after about 50,000 BP, ever more refined and specialized flint tools were made by the Neanderthals and the immigrant Cro-Magnons (knives, blades, skimmers). In this period, they also started to make tools out of bone.
The "modern man" debate and the Great Leap Forward
Until about 50,000–40,000 years ago, the use of stone tools seems to have progressed stepwise: Each phase (habilis, ergaster, and neanderthal) started at a higher level than the previous one, but once that phase had started, further development was slow. In other words, one might call these Homo species culturally conservative.
After 50,000 BP, in what Jared Diamond, author of The Third Chimpanzee, and other anthropologists characterize as a "Great Leap Forward," human culture apparently started to change at much greater speed: "Modern" humans started to bury their dead carefully, made clothing out of hides, developed sophisticated hunting techniques (such as pitfall traps, or driving animals to fall off cliffs), and made cave paintings. This speed-up of cultural change seems connected with the arrival of modern humans, Homo sapiens sapiens.
Additionally, human culture began to become more technologically advanced, in that different populations of humans begin to create novelty in existing technologies. Artifacts such as fish hooks, buttons, and bone needles begin to show signs of variation among different population of humans, something what had not been seen in human cultures prior to 50,000 BP.
Theoretically, modern human behavior is taken to include four ingredient capabilities: Abstract thinking (concepts free from specific examples), planning (taking steps to achieve a farther goal), innovation (finding new solutions), and symbolic behavior (such as images, or rituals). Among concrete examples of modern human behavior, anthropologists include specialization of tools, use of jewelry and images (such as cave drawings), organization of living space, rituals (for example, burials with grave gifts), specialized hunting techniques, exploration of less hospitable geographical areas, and barter trade networks. Debate continues whether there was indeed a "Revolution" leading to modern humanity ("the big bang of human consciousness"), or a more gradual evolution.
Species list
This list is in chronological order across the page by genus.
|
|
|
See also
ReferencesISBN links support NWE through referral fees
- Argue, D., D. Donlon, C. Groves, and R. Wright. 2006. Homo floresiensis: Microcephalic, pygmoid, Australopithecus, or Homo? J. Hum. Evol. 51(4): 360–74. PMID 16919706.
- Banks, W. E, F. d'Errico, A. Townsend Peterson, M. Kageyama, A. Sima, and M.-F. Sánchez-Goñi. 2008. Neanderthal extinction by competitive exclusion. PLoS ONE 3(12): e3972. PMID 19107186.
- Brown, P., T. Sutikna, M. J. Morwood, et al. 2004. A new small-bodied hominin from the Late Pleistocene of Flores, Indonesia. Nature 431(7012): 1055–61. PMID 15514638.
- Callaway, E. 2011. First Aboriginal genome sequenced. Nature News September 22, 2011.
- Callaway, E. 2013. Hominin DNA baffles experts. Nature 504: 16-17. Retrieved December 19, 2013.
- Cann, R. L., M. Stoneking, and A. C. Wilson. 1987. Mitochondrial DNA and human evolution. Nature 325: 31-36.
- Chimpanzee Sequencing and Analysis Consortium. 2005. Initial sequence of the chimpanzee genome and comparison with the human genome. Nature 437:69–87.
- Dartmouth Undergraduate Journal of Science (DUJS). 2013. High-quality archaic human genome sequencing. Dartmouth Undergraduate Journal of Science March 10, 2013. Retrieved December 18, 2013.
- Dawkins, R. 2004. The Ancestor's Tale: A Pilgrimage to the Dawn of Life. Weidenfeld & Nicolson. ISBN 0-7528-7321-0
- Ebersberger, I., D. Metzler, C. Schwarz, and S. Paabo. 2002. Genomewide comparison of DNA sequences between humans and chimpanzees. American Journal of Human Genetics 70:1490–97.
- Fagundes, N. J. R., N. Ray, M. Beaumont, S. Neuenschwander, F. M. Salzano, S. L. Bonatto, and L. Excoffier. 2007. Statistical evaluation of alternative models of human evolution. Proceedings of the National Academy of Sciences 104(45): 17614–17619.
- Gibbons, A. 2011. Who were the Denisovans? Science 333: 1084-1087. Retrieved December 19, 2013.
- Gibbons, A. 2013. Elusive Denisovans sighted in oldest human DNA. Science 342(6163): 1156.
- Gould, S. J. 1990. Men of the Thirty-third Division. Natural History April, 1990: 12,14,16-18, 20, 22-24.
- Gould, S. J. 1994. Lucy on the Earth in stasis. Natural History September, 1994: 12,14,16-18, 20.
- Green, R. E., J. Krause, A. W. Briggs, T. Maricic, et al. 2010. A draft sequence of the Neanderthal genome. Science 328(5979): 710-722.
- Green, R.E., A.-S. Malaspinas, J. Krause, et al. 2008. A complete Neanderthal mitochondrial genome sequence determined by high-throughput sequencing. Cell 134(3): 416–426. PMID 18692465.
- Gutiérrez, G. D. Sánchez, and A. Marín. 2002. A reanalysis of the ancient mitochondrial DNA sequences recovered from Neandertal bones. Mol. Biol. Evol. 19(8): 1359–66. PMID 12140248.
- Haldane, J. B. S. 1955. Origin of man. Nature 176: 169-170.
- Hammer, M. F., a. E. Woerner, F. L. Mendez, J. C. Watkins, and J. D. Wall. 2011. Genetic evidence for archaic admixture in Africa. Proceedings of the National Academy of Sciences 108(37): 15123–15128.
- Harvati,K. 2003. The Neanderthal taxonomic position: Models of intra- and inter-specific craniofacial variation. J. Hum. Evol. 44(1): 107–32. PMID 12604307.
- Hebsgaard, M. B., C. Wiuf, M. T. Gilbert, H. Glenner, and E. Willerslev. 2007. Evaluating Neanderthal genetics and phylogeny. J. Mol. Evol. 64(1): 50–60. PMID 17146600.
- Heizmann, E. P. J. and D. R. Begun. 2001. The oldest Eurasian hominoid. Journal of Human Evolution 41(5): 463-481.
- Hodgson, J. A., and T. R. Disotell. 2008. No evidence of a Neanderthal contribution to modern human diversity. Genome Biology 9(2): 206.
- Hortolà, P. and B. Martínez-Navarro. 2012. The Quaternary megafaunal extinction and the fate of Neanderthals: An integrative working hypothesis. Quaternary International
- Ijdo, J. W., A. Baldini, D. C. Ward, S. T. Reeders, and R. A. Wells. 1991. Origin of human chromosome 2: An ancestral telomere-telomere fusion. Genetics 88: 9051-9055. Retrieved February 9, 2014.
- Indiana University (IU). 2006. Scientists discover hominid cranium in Ethiopia. Indiana University News, March 27, 2006. Retrieved February 9, 2014.
- International Chimpanzee Chromosome 22 Consortium. 2004. DNA sequence and comparative analysis of chimpanzee chromosome 22. Nature 429: 382–88.
- Kordos, L., and D. R. Begun. 2001. Primates from Rudabánya: allocation of specimens to individuals, sex and age categories. Journal of Human Evolution 40(1): 17-40.
- Kreger, C. D. 2005. Homo sapiens: Introduction. Archaeology.info.
- Krings, M., A. Stone, R. W. Schmitz, H. Krainitzki, M. Stoneking, and S. Pääbo. 1997. Neandertal DNA sequences and the origin of modern humans. Cell 90(1): 19-30. PMID 9230299.
- Lordkipanidze, D., M. S. Ponce de León, A. Margvelashvili, et al. 2013. A complete skull from Dmanisi, Georgia, and the evolutionary biology of early Homo. Science 342(6156): 326-331. Retrieved October 16, 2013.
- Liu, G., NISC Comparative Sequencing Program, S. Zhao, J. A. Bailey, S. C. Sahinalp, C. Alkan, E. Tuzun, E. D. Green, and E. E. Eichler. 2003. Analysis of primate genomic variation reveals a repeat-driven expansion of the human genome. Genome Research 13:358–68.
- Marshall, M. 2013. Mystery human species emerges from Denisovan genome. New Scientist November 19, 2013. Retrieved December 16, 2013.
- Martin, R. D., A. M. Maclarnon, J. L. Phillips, and W. B. Dobyns. 2006. Flores hominid: New species or microcephalic dwarf? The Anatomical Record. Part A, Discoveries in Molecular, cellular, and Evolutionary Biology 288(11): 1123–45. PMID 17031806.
- Mason, P. H., and R. V. Short. 2011. Neanderthal-human hybrids. Hypothesis 9(1): e1.
- Mayr, E. 2001. What Evolution Is. New York: Basic Books. ISBN 0465044255
- Mcbrearty, S., and A. S. Brooks. 2000. The revolution that wasn't: A new interpretation of the origin of modern human behavior. "Journal of Human Evolution" 39(5): 453–563. PMID 11102266.
- Meyer, M., M. Kircher, M. T. Gansauge, et al. 2012. A high-coverage genome sequence from an archaic Denisovan individual. Science 338(6104): 222-226. Retrieved October 19, 2013.
- Mellars, P. 2006. Why did modern human populations disperse from Africa ca. 60,000 years ago?. "Proceedings of the National Academy of Sciences" 103 (25): 9381–6. PMID 16772383. Retrieved October 19,2013.
- Mellars, P., and J. C. French. 2011. Tenfold population increase in Western Europe at the Neandertal – to – modern human transition. Science 333(6042): 623–627. PMID 21798948.
- National Geographic (NG). 2013. Why am I denisovan. The Genographic Project. Retrieved October 16, 2013.
- Nielsen, R., C. Bustamante, A. G. Clark, S. Glanowski, T. B. Sackton, M. J. Hubisz, A. Fiedel-Alon, D. M. Tanenbaum, D. Civello, T. J. White, J. J. Sninsky, M. D. Adams, and M. Cargill. 2005. A scan for positively selected genes in the genomes of humans and chimpanzees. PLoS Biology 3(6):e170.
- O'Neil, D. 2013. Evolution's past is modern human's present. "Behavioral Sciences Department", Palomar College, San Marcos, California. Retrieved December 19, 2013.
- Ovchinnikov, I. V., A. Götherström, G. P. Romanova, V. M. Kharitonov, K. Lidén, and W. Goodwin. 2000. Molecular analysis of Neanderthal DNA from the Northern Caucasus. Nature 404: 490.
- Patterson, N., D. J. Richter, S. Gnerre, E. S. Lander, and D. Reich. 2006. Genetic evidence for complex speciation of humans and chimpanzees. Nature 441: 1103-1108.
- Pennisi, E. 2013. More genomes from Denisova Cave show mixing of early human groups. "Science" 340: 799. Retrieved December 19, 2013.
- Pennsylvania State University (PSU). 1997. DNA shows neandertals were not our ancestors]. Penn State News 1997.
- Rasmussen, M.,X. Guo, Y.Wang, et al. 2011. An Aboriginal Australian genome reveals separate human dispersals into Asia. Science 334(6052): 94-98. Retrieved December 19, 2013.
- Reich, D., R. E. Green, M. Kircher, J. Krause, et al. 2010. Genetic history of an archaic hominin group from Denisova Cave in Siberia. "Nature" 468(7327): 1053–1060. PMID 21179161. Retrieved February 9, 2014.
- Reich, D., N. Patterson, M. Kircher, et. al. 2011. Denisova admixture and the first modern human dispersals into Southeast Asia and Oceania. The American Journal of Human Genetics 89: 516-28. PMID 21944045. Retrieved December 19, 2013.
- Rohde, D. L. T. 2005. On the common ancestors of all living humans. Submitted to American Journal of Physical Anthropology.
- Rohde, D. L. T., S. Olson, and J. T. Chang. 2004. Modelling the recent common ancestry of all living humans. Nature 431: 562-566.
- Scientific American (SA). 2005. Fossil reanalysis pushes back origin of Homo sapiens. "Scientific American " February 17, 2005. Retrieved December 19, 2013.
- Serre, D., A. Langaney, M. Chech, et al. 2004. No evidence of Neandertal mtDNA contribution to early modern humans. PLoS Biol. 2(3): E57. PMID 15024415.
- Smithsonian Institution, Human Origins Program. 2007a. Homo sapiens. Smithsonian National Museum of Natural History. Retrieved February 9, 2014.
- Smithsonian Institution, Human Origins Program. 2007b. Homo erectus. Smithsonian National Museum of Natural History. Retrieved February 9, 2014.
- Stringer, c. 2012. Evolution: What makes a modern human. Nature 485(7396): 33–35.
- Viegas, J. 2010. Toothy tree-swinger may be earliest human. News.discovery.com May 21, 2010. Retrieved February 6, 2014.
- Wall, J. d., and M. F. Hammer. 2006. Archaic admixture in the human genome. Current Opinion in Genetics & Development 16(6): 606–610.
- Wang, C. c., S. E. Farina, and H. Li. 2013. Neanderthal DNA and modern human origins. Quaternary International 295: 126–129.
- Watson, T. 2013. Skull discovery sheds light on human species. USA Today October 17, 2013. Retrieved December 16, 2013.
- Wells, J. 2000. Icons of Evolution. Washington, DC: Regnery Publishing. ISBN 0895262762.
- Wildman, D. E., M. Uddin, G. Liu, L. I. Grossman, and M. Goodman. 2003. Implications of natural selection in shaping 99.4% nonsynonymous DNA identity between humans and chimpanzees: Enlarging genus Homo. Proceedings of the National Academy of Sciences 100:7181–88.
- Wilford, J. N. 2013. Skull fossil suggests simpler human lineage.] New York Times October 17, 2013. Retrieved December 16, 2013.
- Wolfgang, E., et al. 2002. Molecular evolution of FOXP2, a gene involved in speech and language. Nature 418: 870.
- Wood, T. C. 2006. The chimpanzee genome and the problem of biological similarity]. Occassional Papers of the BSG 7:1–18.
- Wood, B., and M. Collard. 1999. The changing face of genus Homo. Evol. Anth. 8(6): 195-207.
- Wood, B. and B. G. Richmond. 2000. Human evolution: Taxonomy and paleobiology. Journal of Anatomy 197 (Pt 1): 19–60. PMID 10999270. Retrieved December 19, 2013.
- Yunis, J. J., and O. Prakash. 1982. The origin of man: A chromosomal pictorial legacy. Science 215: 1525–30.
- Zimmer, C. 2010. Denisovans were Neanderthals' cousins, DNA analysis reveals. New York Times December 22, 2010.
- Zimmer, C. 2013. Toe fossil provides complete Neanderthal genome. New York Times December 18, 2013.
Credits
New World Encyclopedia writers and editors rewrote and completed the Wikipedia article in accordance with New World Encyclopedia standards. This article abides by terms of the Creative Commons CC-by-sa 3.0 License (CC-by-sa), which may be used and disseminated with proper attribution. Credit is due under the terms of this license that can reference both the New World Encyclopedia contributors and the selfless volunteer contributors of the Wikimedia Foundation. To cite this article click here for a list of acceptable citing formats.The history of earlier contributions by wikipedians is accessible to researchers here:
- Human_evolution history
- Cro-Magnon history
- Mitochondrial_Eve history
- Archaic_human_admixture_with_Homo_sapiens history
The history of this article since it was imported to New World Encyclopedia:
Note: Some restrictions may apply to use of individual images which are separately licensed.