Glial cell


Glial cells, commonly called neuroglia or simply glia, are one of two major classes of cells in neural tissues, the other being neurons, for which the glial cells provide support. Glial cells surround neurons, hold them in place, provide nutrition (nutrients and oxygen), help maintain homeostasis, provide electrical insulation, destroy pathogens, regulate neuronal repair and the removal dead neurons, and participate in signal transmission in the nervous system. They help in guiding the construction of the nervous system and control the chemical and ionic environment of the neurons (Alberts et al. 1989). In the mammalian brain, glial cells are estimated to outnumber neurons by about ten to one (Alberts et al. 1989).
Glia, which is Greek for glue, have traditionally been viewed as the glue that holds the neuronal system together. However, there is increasing evidence that the type of glia known as astrocytes may play a more active role in brain function, by influencing and perhaps directing the activity of neurons (SFN 2000).
In a living organism, cells depend upon other cells to perform their particular functions, to allow the smooth functioning of tissues, organs, organ systems, and the rest of the body. In the case of nervous tissue, while neurons have traditionally been seen as being in the "starring" role and glial cells in a "supporting" role, both are essential and work together harmoniously for the correct functioning of the central nervous system and the peripheral nervous system.
Overview

There are two main classes of cells in neural tissue: Neurons and glial cells, with neurons historically having been identified as being primary in function and the glial cells having been identified as being secondary and supportive in function. Newer studies are revealing a much more balanced sharing of the nervous system function between neurons and glial cells. In the traditional model, Neurons (also known as neurones and nerve cells) are electrically excitable cells that process and transmit signals from both internal and external environments. The process by which the mind converts those signals to information is still the subject of study and debate.
In vertebrates, neurons are the core components of the brain, spinal cord, and peripheral nerves. Although the neuron is considered a discrete unit, the output of the nervous system is produced by the connectivity of neurons (that is, the strength and configuration of the connections between neurons). The basic function of a neuron, then, is to process signals as part of a larger complex of neurons, each neuron receiving signals from several sources (usually neurons), modifying the signal and passing it on to several receivers (usually neurons). Signal transfer between neurons is achieved via chemical or electric impulses across a synapse (the junction between cells).
Glial cells in the traditional model are typically described as having a supporting role to the neurons. The glial cells surround both the central cell bodies and the extensions of neurons and occupy the space between neurons (Alberts et al. 1989). In the vertebrate peripheral nervous system, the best understood glial cells are the Schwann cells, while in the vertebrate central nervous system the best understood glial cells are the oligodendrocytes. Both of these provide electrical insulation by wrapping themselves around axons in the form of a myelin sheath (Alberts et al. 1989). Besides the oligodendrocytes, the other three main types of glial cells in the central nervous system are microglia cells, which are involved in reactions to tissue damage and infection; ependymal cells, which form the ciliated epithelial lining of the central cavity and often have processes ending in blood vessels; and astrocytes, which are the most plentiful and have many radiating processes (Alberts et al. 1989). Some of these later processes end on the surface of neurons, some form the external surface layer of the central nervous system and surround blood vessels, collaborating with endothelial cells of the blood capillaries to help create the blood-brain barrier.
Function of glial cells
Some glia function primarily as physical support for neurons. Others regulate the internal environment of the brain, especially the fluid surrounding neurons and their synapses, and provide nutrition to nerve cells. Glia have important developmental roles, guiding migration of neurons in early development, and producing molecules that modify the growth of axons and dendrites. Recent findings in the hippocampus and cerebellum have indicated that glia are also active participants in synaptic transmission, regulating clearance of neurotransmitter from the synaptic cleft, releasing factors, such as ATP, that modulate presynaptic function, and even releasing neurotransmitters themselves.
Traditionally, glia had been thought to lack certain features possessed by neurons. For example, glia were not believed to have chemical synapses or to release neurotransmitters. They were considered to be the passive bystanders of neural transmission. However, recent studies have countered this assertion.
For example, astrocytes are crucial in clearance of neurotransmitters from within the synaptic cleft, which provides distinction between arrival of action potentials and prevents toxic build up of certain neurotransmitters, such as glutamate (excitotoxicity). Furthermore, at least in vitro, astrocytes can release the neurotransmitter glutamate in response to certain stimulation. Another unique type of glia, the oligodendrocyte precursor cells or OPCs, have very well defined and functional synapses from at least two major groups of neurons. The only notable differences between neurons and glia, by modern scrutiny, are the ability to generate action potentials and the polarity of neurons, namely the axons and dendrites, which glia lack. It is inappropriate in modern contexts to consider glia simply as "glue" in the nervous system, as the name implies. They are also crucial in the development of the nervous system and in processes such as synaptic plasticity and synaptogenesis.
Glia have a role in the regulation of repair of neurons after injury. In the central nervous system (CNS), glia suppress repair. Astrocytes enlarge and proliferate to form a scar and produce myelin and inhibitory molecules that inhibit regrowth of a damaged or severed axon. In the peripheral nervous system (PNS), Schwann cells promote repair. After axon injury, Schwann cells regress to an earlier developmental state to encourage regrowth of the axon. This difference between PNS and CNS raises hopes for the regeneration of nervous tissue in the CNS, for example, with a spinal cord injury or severance.
Types of glia
Microglia
Microglia are specialized macrophages, capable of phagocytosis, that protect neurons of the central nervous system. Though not technically glia because they are derived from hemopoietic precursors rather than ectodermal tissue, they are commonly categorized as such because of their supportive role to neurons.
These cells comprise approximately 15 percent of the total cells of the central nervous system. They are found in all regions of the brain and spinal cord. Microglial cells are small relative to macroglial cells, with changing shapes and oblong nuclei. They are mobile within the brain and multiply when the brain is damaged. In the healthy central nervous system, microglia processes constantly sample all aspects of their environment (neurons, macroglia, and blood vessels).
Macroglia
| Location | Name | Description |
| CNS | Astrocytes |
The most abundant type of glial cell, astrocytes (also called astroglia) have numerous projections that anchor neurons to their blood supply. They regulate the external chemical environment of neurons by removing excess ions, notably potassium, and recycling neurotransmitters released during synaptic transmission. Current theories suggest that astrocytes may be the predominant "building blocks" of the blood-brain barrier. Astrocytes may regulate vasoconstriction and vasodilation by producing substances such as arachidonic acid, which yields vasoactive metabolites. Astrocytes signal each other using calcium. The gap junctions (also known as electrical synapses) between astrocytes allow the messenger molecule IP3 to diffuse from one astrocyte to another. IP3 activates calcium channels on cellular organelles, releasing calcium into the cytoplasm. This calcium may stimulate the production of more IP3. The net effect is a calcium wave that propagates from cell to cell. Extracellular release of ATP, and consequent activation of purinergic receptors on other astrocytes, may also mediate calcium waves in some cases. The two general types of astrocytes, protoplasmic and fibrous, are similar in function but distinct in morphology and distribution. Protoplasmic astrocytes have short, thick, highly branched processes and are typically found in gray matter. Fibrous astrocytes have long, thin, less branched processes and are more commonly found in white matter. |
| CNS | Oligodendrocytes |
Oligodendrocytes are cells that coat axons in the central nervous system (CNS) with their cell membrane, called myelin, producing the so-called myelin sheath. The myelin sheath provides insulation to the axon, which enhances the efficiency of [electricity|electrical]] signal propagation along the axon. |
| CNS | Ependymal cells |
Ependymal cells, also named ependymocytes, form the surface layer of the wall of the system of interconnected, fluid-filled cavities of the brain and spinal cord. These cells create and secrete cerebrospinal fluid (CSF) and beat their cilia to help circulate it. |
| CNS | Radial glia |
Radial glia cells arise from neuroepithelial cells after the onset of neurogenesis, but their differentiation abilities are more restricted than those of neuroepithelial cells. In the developing nervous system, radial glia function both as neuronal progenitors and as a scaffold upon which newborn neurons migrate. In mature vertebrates, the cerebellum in the brain and retina in the eyes retain characteristic radial glial cells. In the cerebellum, these are Bergmann glia, which regulate synaptic plasticity and in the retina, the principal glial cell is the radial Müller cell, which participates in a bidirectional communication with neurons. |
| PNS | Schwann cells |
Similar in function to oligodendrocytes, Schwann cells provide myelination to axons in the peripheral nervous system (PNS). They also have phagocytotic activity and clear cellular debris in support of the regrowth of PNS neurons. |
| PNS | Satellite cells |
Satellite cells are small cells that line the exterior surface of PNS neurons and help regulate the external chemical environment. |
Capacity to divide
Glia retain the ability to undergo cell division in adulthood, while most neurons cannot. The view is based on the general deficiency of the mature nervous system in replacing neurons after an insult or injury, such as a stroke or trauma, while very often there is a profound proliferation of glia, or gliosis, near or at the site of damage. However, detailed studies found no evidence that "mature glia," such as astrocytes or oligodendrocytes, retain the ability of mitosis. Only the resident oligodendrocyte precursor cells seem to keep this ability after the nervous system matures. On the other hand, there are a few regions in the mature nervous system, such as the dentate gyrus of the hippocampus and the subventricular zone, where generation of new neurons has been observed.
Embryological development
Most glia are derived from ectodermal tissue of the developing embryo, particularly the neural tube and crest. The exception is microglia, which are derived from hemopoietic stem cells. In the adult, microglia are largely a self-renewing population that is distinct from macrophages and monocytes, which infiltrate the injured and diseased central nervous system.
In the central nervous system, glia develop from the ventricular zone of the neural tube. These glia include the oligodendrocytes, ependymal cells, and astrocytes. In the peripheral nervous system, glia derive from the neural crest. These PNS glia include Schwann cells, in nerves, and satellite cells, in ganglia.
History
Glia were discovered in 1856, by the pathologist Rudolf Virchow in his search for a "connective tissue" in the brain.
The human brain contains about ten times more glial cells than neurons (SFN 2000). Following its discovery in the late nineteenth century, this fact underwent significant media distortion, emerging as the famous myth claiming that people "are using only 10 percent of their brain." The role of glial cells as managers of communications in the synapse gap, thus modifying the pace of learning, has been discovered only very recently (2004).
Additional images
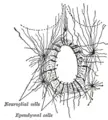
Section of central canal of medulla spinalis, showing ependymal and neuroglial cells.
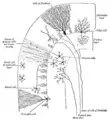
Transverse section of a cerebellar folium.


ReferencesISBN links support NWE through referral fees
- Alberts, B., D. Bray, J. Lewis, M. Raff, K. Roberts, and J. D. Watson. 1989. Molecular Biology of the Cell. New York: Garland Publishing. ISBN 0824036956.
- Overstreet, L.S. 2005. Quantal transmission: not just for neurons. Trends Neurosci 28(2): 59-62. Retrieved September 22, 2007.
- Peters, A. 2004. A fourth type of neuroglial cell in the adult central nervous system. J Neurocytol 33(3): 345-357.
- Society for Neuroscience (SFN). 2000. Astrocytes. Brain Briefings. December 2000.
- Than, K. 2006. New source of replacement brain cells found. LiveScience. Retrieved September 22, 2007.
- Volterra, A., and C. Steinhäuser. 2004. Glial modulation of synaptic transmission in the hippocampus. Glia 47(3): 249-257.
Credits
New World Encyclopedia writers and editors rewrote and completed the Wikipedia article in accordance with New World Encyclopedia standards. This article abides by terms of the Creative Commons CC-by-sa 3.0 License (CC-by-sa), which may be used and disseminated with proper attribution. Credit is due under the terms of this license that can reference both the New World Encyclopedia contributors and the selfless volunteer contributors of the Wikimedia Foundation. To cite this article click here for a list of acceptable citing formats.The history of earlier contributions by wikipedians is accessible to researchers here:
The history of this article since it was imported to New World Encyclopedia:
Note: Some restrictions may apply to use of individual images which are separately licensed.